В спинном мозгу ноцицептивные афференты оканчиваются на нейронах в заднем роге. Эти клетки дают начало трактам переднебоковых канатиков (сшгаоталамическим и др.), подробно описанным в предыдущей главе. Они восходят к стволу мозга, где к ним присоединяются ноцицептивные афференты от области головы (в основном из состава тройничного нерва); затем информация направляется к таламусу (рис. 10.7, слева; см. также рис. 9.20). Сведения о механизме работы этих восходящих систем, их последующих центростремительных проекциях и об участии ретикулярной формации, таламуса и коры больших полушарий в переработке ноцицептивных сигналов приводятся в соответствующих разделах предыдущей главы [14, 30].
Здесь мы остановимся лишь на участии коры больших полушарий в ноцицепции и формировании болевых ощущений,в частности на его сенсорно-дискриминативном и когнитивном компонентах, только ради того, чтобы упомянуть о господствовавшем в первой половине нашего столетия, основанном на клинических и экспериментальных данных мнении, согласно которому кора для проявления таких сознательных ощущений вовсе не обязательна. Главным центром их возникновения считался таламус. Последующие тщательные наблюдения, особенно над людьми, получившими мозговые травмы во время второй мировой войны, привели к пересмотру этой точки зрения. Было установлено, что повреждение определенных корковых областей в глу-
ГЛАВА 10. НОЦИЦЕПЦИЯ И БОЛЬ 229
|
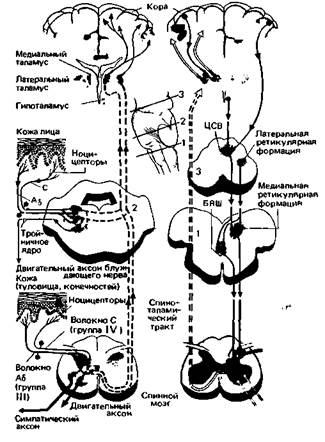
| Рис. 10.7. Схема, демонстрирующая восходящие ноцицептивные пути {слева) и системы нисходящих трактов, модулирующих ноцицептивные сигналы (справа). Из восходящих систем показаны только спиноталамический тракт и соединяющиеся с ним тройничноталамические пути. Другие тракты, участвующие в восходящем проведении ноцицептивной информации (например, спиноретикулярный и спиноцервикальный), опущены для простоты изображения. Специфичные таламокортикальные тракты начинаются в латеральном таламусе и оканчиваются главным образом в соматосенсорной коре. Эфференты медиальных таламических ядер более диффузны; они проецируются не только на обширные области лобной коры, но и на подкорковые структуры, особенно на лимбическую систему (она сама и ретикулярные входы в эти ядра не показаны). Нисходящие системы действуют преимущественно на спинальном уровне (или на не показанные здесь соответствующие тройничные структуры). Центральная вреэко-вид сбоку ствола мозга с уровнями разреза: 1 -краниальный край нижней оливы; 2 середина моста; 3 нижняя часть среднего мозга; ЦСВ центральное серое вещество среднего мозга; БЯШ-большое ядро шва (nucleus raphe magnus) (по экспериментальным данным многих авторов)
|
бине теменной части центральной борозды вызывает устойчивую контралатеральную нечувствитель-
ность к боли. В некоторых случаях она только частичная (например, у руки или ноги), в других затрагивается вся половина тела. Позднее удалось подтвердить такие данные экспериментально: локальная электрическая стимуляция этих зон головного мозга вызывает у людей боль. Значит, можно сделать вывод, что болевые ощущения, как и все прочие сознательные сенсорные процессы, невозможны без коры больших полушарий [29].




