Переработка информации в нейронах соматосенсорной коры
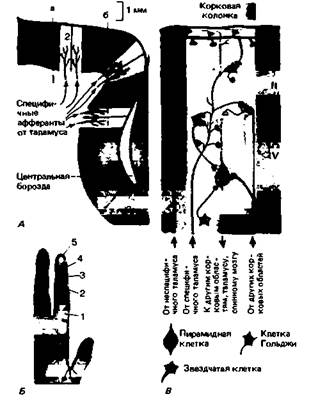
Колонки корковых нейронов. При прохождении микроэлектрода перпендикулярно поверхности постцентральной извилины (линия а на рис. 9.25, А) последовательно встречаемые им нейроны часто обладают идентичными или широко перекрывающимися рецептивными полями (рис. 9.25, Б). Если же электрод перемещается под утлом к этой поверхности (линия б), он проходит нейроны с соседними, но четко разграниченными рецептивными полями (в соответствии с соматотопической организацией). Из этих и других данных сделан вывод, что соматосен-
сорная кора, как и двигательная (см. гл. 5, с. 121), организована в виде функциональных единиц - колонок нейронов, перпендикулярных ее поверхности; на рис. 9.25, А их схематически изображено шесть. Эти колонки диаметром 0,2-0,5 мм отражают два анатомических свойства коры-ограниченное горизонтальное распространение окончаний афферентных нейронов, идущих от вентробазального таламического ядра, и предпочтительно вертикальную ориентацию дендритов пирамидных клеток (рис. 9.25, В). У мышей колонки, соответствующие вибриссам, особенно развиты; из-за своего внешнего 218 ЧАСТЬ III. ОБЩАЯ И СПЕЦИАЛЬНАЯ СЕНСОРНАЯ ФИЗИОЛОГИЯ вида, выявляемого при гистологическом окрашивании, они названы «бочонками». Рецепторная специфичность колонок. Адекватная избирательная стимуляция, например, различных кожных рецепторов (МА, БА и ТП; см. разд. 9.2) показывает, что нейроны одной колонки возбуждаются обычно рецепторами только одного типа. Очевидно, колонки - это функциональные единицы, соответствующие положению и сенсорной специфике периферических сенсорных нервных окончаний. В SI и SII обнаружены также терморецептивные и ноцицептивные нейроны, хотя они менее (на 1-2 порядка) многочисленны, чем механорецептивные. Собраны ли они в колонки, неясно. Возможно, обработка информации в колонках и между ними по своей природе иерархическая. Эта гипотеза подтверждается экспериментальными наблюдениями, предполагающими разделение корковых нейронов на простые («низшие») и сложные («высшие»). Такое представление наиболее подробно разработано для зрительной системы (см. гл. 11). Простые и сложные нейроны. Характеристики импульсации простых корковых нейронов очень близки к таковым у связанных с ними рецепторов. Например, в SI есть нейроны, ведущие себя как рецепторы БА. Корковые нейроны, отвечающие на периферические стимулы импульсацией, явно отличающейся от той, что свойственна связанным с ними рецепторам, называются сложными. Этот термин охватывает разнообразные клетки. Среди них известны нейроны, которые реагируют только на стимулы, движущиеся линейно по поверхности кожи, и дают максимальный ответ на движение в определенном направлении. Они обнаружены в SI, Sil и ассоциативных областях теменной коры (поля 5 и 7 Бродмана) [3]. Терморецептивные корковые нейроны,найденные у обезьяны, относятся к сложным. В отличие от периферических терморецепторов (разд. 9.4) они реагируют либо только на изменения температур кожи, либо только на ее постоянную температуру. Кроме того, у них нет пика ответа при температуре около 26 "С, характерного для холодовых рецепторов (рис. 9.12). Другой тип сложных терморецепторных нейронов обнаружен в коре мозга крысы. Здесь частота импульсации «скачет» между максимумом и минимумом, когда температура соответствующих рецептивных полей кожи меняется приблизительно на 2 °С (пороговый детектор [7]). Эти примеры простых и сложных нейронов показывают, что корковая переработка сигналов приводит к нейронному представительству или отфильтровыванию информации о самых разнообразных параметрах периферических стимулов. Такая способность извлекать информацию, названная выделением признаков [2, 23], особенно тщательно изучена в зрительной коре (см. гл. 11, с. 258). Эфферентные связи SI. От области S1, как и от всех прочих участков коры, отходит множество эфферентных аксонов (рис. 9.25, В). Как полагают, они проводят в переработанном виде информацию о периферических стимулах к другим частям центральной нервной системы. SI связана со следующими областями (указана и основная функция связи): -двигательная кора: регуляция движения по обратной связи; -теменные ассоциативные зоны: интеграция зрительной и тактильной информации; —контралатеральные SI и Sil: интеграция билатеральной тактильной информации; -таламус, ядра заднего столба, спинной мозг: эфферентная регуляция потока афферентной информации (разд. 9.12, с. 219). Корковая область SII значительно меньше SI. Она лежит у латерального конца постцентральной извилины, на верхней стенке сильвиевой борозды. Здесь соматотопическое представительство поверхности тела билатеральное. Рецептивные поля нейронных колонок обычно находятся на обеих сторонах тела, а их проекции часто расположены симметрично. Как полагают, SII играет особую роль в сенсорной и двигательной координации двух сторон тела (например, при хватании или ощупывании обеими руками) [2].
|