Из ботанических групп растений преобладающее большинство составляли двудольные виды сорных растений. Однодольные виды были представлены растениями семейства Poaceae (табл.1).
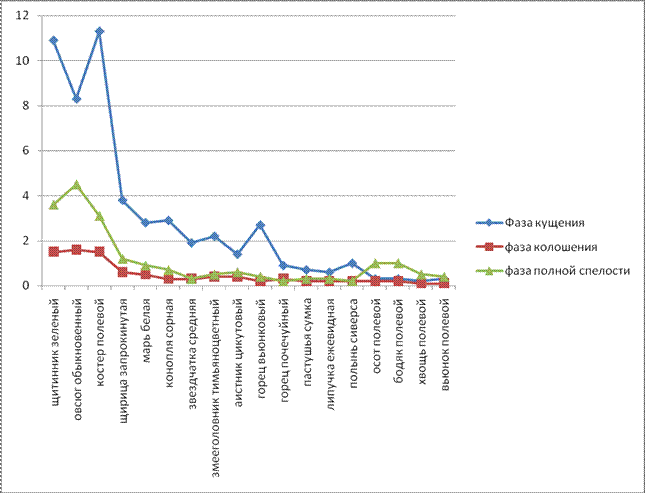
Сезонная динамика сорного компонента во многом определяется метеорологическими условиями. Засоренность посева на контрольном варианте (без обработки) опыта в фазу кущения пшеницы вирировала в пределах 52,5- 63,2 шт/м2, с преобладанием по численности малолетних видов (табл.2). Из мнологолетних двудольных преобладали осот полевой, и бодяг полевой в количестве 2,3 шт/м2 . Из малолетних однодольных наибольшим количеством наблюдались Щетинник зеленый (16,5 шт/м2) и костер полевой (16,3 шт/м2), наименьшим количеством овсюг обыкновенный (13,9 шт/м2). Из малолетних двудольных преобладали Щирица запрокинутая (5,2 шт/м2) и аистник цикутовый (2,3 шт/м2). В фазу колошения яровой пшеницы на контрольном варианте численность многолетних двудольных сорняков была такой: хвощ полевой и вьюнок полевой (0,5 шт/м2); из малолетних однодольных овсюг обыкновенный (2,4 шт/м2); и из малолетних двудольных щирица запрокинутая (0,9 шт/м2).
Рис. 1.Соотношение ботанических классов сорных растений в пшеничном агрофитоценозе за годы исследований.
В фазу спелости яровой пшеницы на контрольном варианте численность многолетних сорняков уменьшилась на 2,78 %. (табл. 2). Дополнительное отрастание розеток Sonchus arvensis связано с обильными осадками в первой декаде июля и оптимальной температурой для их прорастания (15,80С). По мнению Б.М. Смирнова (1958), к этому времени наступает второй период их жизни - интенсивное образование новых горизонтальных корней, что увеличивает площадь засорения. Численность малолетних видов увеличилась за счет появления в посеве Щетинник зеленый, Овсюг обыкновенный, Костер Полевой.
К концу периода вегетации такие сорные растения, как пастушья сумка, липучка ежевидная Lappula squarroosa исчезают из агрофитоценозе. Это связанно с их биологическими особенностями, так как они прорастают рано весной и к периоду полной спелости яровой пшеницы созревают, плодоносят и заканчивают свой жизненный цикл.
Табл.2
Количественный состав сорных растений пшеничного агрофитоценозе на контрольном варианте (без обработки), шт./м2 2009 г.
Из таблицы 3 видно, что в год исследования наибольшее развитие получили ранние и поздние яровые сорняки, их количества в фазу кущения яровой пшеницы были максимальным среди других биологических групп. Из этого видно что, численность сорняков увеличилась в фазе кущения и уменьшилась к фазе колошения яровой пшеницы. Эта особенность объясняется биологией сорных растений. В фазу кущения яровой пшеницы данные сорные растения находятся на начальном этапе развития. В фазу колошения яровой пшеницы они цветут и плодоносят, а к моменту спелости пшеницы большинство из них отмирает.
Многолетние сорные растения Вьюнок полевой Convolvulus arvensis, Хвощ полевой Eguisetum arvense на протяжении периода вегетации слабо изменяли свою численность. Они постепенно проходили все этапы своего развития.
Засоренность посева на комбинированной обработке в фазу кущения пшеницы менялась в пределах 0,6-58,2 шт/м2, с преобладанием по численности малолетних однодольных видов (табл. 3).
В фазу колошения яровой пшеницы на комбинированной обработке численность многолетних сорняков уменьшилась на 22%, а малолетних однодольных и двудольных увеличилась на 77% (табл. 3). Из многолетних сорняков численность Осота полевого (Sonchus arvensis) уменьшилась, а из малолетних сорняков увеличилась за счет появления в посеве Овсюга обыкновенного (Avena fatua) и Мари белой (Chenopodium album).
К концу фазы полной спелости яровой пшеницы такие сорные растения как: Марь белая (Chenopodium album) и Аистник цикутовый (Erodium cicutarium) – практически исчезли из агрофитоциноза. Это связано с биологическими особенностями, так как они прорастают весной, и к периоду спелости яровой пшеницы созревают, плодоносят и заканчивают свой жизненный цикл.
Табл. 3
Количественный состав сорных растений пшеничного агрофитоценозе на комбинированной обработке, шт./м2 2009 г
На мульчирующей разноглубинной обработке в фазу кущения яровой пшеницы произрастало 18 видов сорных растений. Среди них наибольшее распространение получили ранние и поздние яровые: Марь белая (Chenopodium album), Щетинник зеленый (Setaria Beauv), Овсюг обыкновенный (Avena fatua) – 11% и 2 шт/м2.
При минимальном количестве многолетних 22% (табл. 2.8).В фазу колошения яровой пшеницы преобладание ранних и поздних яровых сорняков сохранилась, и уменьшалось количество зимующих до 5%, количество многолетних осталось таким же. К концу периода вегетации яровой пшеницы такие же сорные растения, Марь белая (Chenopodium album) исчез из агрофитоценоза. Это связанно с его биологической особенностью, так как он прорастает весной и к периоду спелости яровой пшеницы созревает, плодоносит и заканчивает свой жизненный цикл.
Рис.2 Видовой состав сорных растений в посевах яровой пшеницы на контроле
(без обработки)
Рис.3 Видовой состав сорных растений в посевах яровой пшеницы на комбинированной обработке
Рис. 4 Видовой состав сорных растений в посевах яровой пшеницы
Количественный состав сорных растений пшеничного агрофитоценозе на мульчирующей разноглубинной обработке, шт./м2 2009 г В фазу колошения яровой пшеницы на контрольном варианте из многолетних сорняков возросла численность Avena fatua и в посеве появилась Setaria Beauv. Дополнительное отрастание розеток Sonchus arvensis связано с обильными осадками в первой декаде июля и оптимальной температурой для их прорастания (15,80С). По мнению Б.М. Смирнова (1958), к этому времени наступает второй период их жизни - интенсивное образование новых горизонтальных корней, что увеличивает площадь засорения. Численность малолетних видов увеличилась за счет появления в посеве Щетинника зеленого, Овсюга обыкновенного, Костера полевого.
К концу периода вегетации такие сорные растения, как пастушья сумка, липучка ежевидная Lappula squarroosa исчезают из агрофитоценозе. Это связанно с их биологическими особенностями, так как они прорастают рано весной и к периоду полной спелости яровой пшеницы созревают, плодоносят и заканчивают свой жизненный цикл.
В 2009 году сегетальная растительность была представлена 18 видами сорняков. При температуре почвы в мае 14,80С наблюдалась массовое пояпление в посеве Avena fatua.
Из многолетних преобладал Осот желтый Sonchus arvensis, среди малолетних доминировали яровые сорняки - Звездчатка средняя Stellaria media, Марь белая Chenopodium album. По численности преобладали малолетние яровые виды (0,2-2,3 шт.⁄м2). На протяжении вегетации культуры численность многолетних сорняков оставалась на одном уровне. Малолетние же сорняки увеличили свою численность за счет появления новых видов и увеличения количества особей существующих видов. По данным Г.Ш.Турсумбековой (2006), увеличение численности зимующих сорняков происходит в засушливых условиях года. Общим для 2009 года исследования было быстрое развитие сорных растений в первую половину вегетации, достижение максимума в фазе колошения яровой пшеницы, а затем постепенно их снижение.
Рис.5. Изменение численности сорных растений в течение вегетации на 2009 г, шт⁄м2.
Погодные условия, как уже отмечалось, во многом определяют массовое появление или исчезновение сорных растений в посевах.
Весной интенсивное появление розеток Sonchus arvensis происходило при прогревании почвы до 13-150С. При 100С (2009 г)их численность была значительно ниже. По данным Б.М. Смирнова (1958), первый период образования побегов происходит при прогревании почвы до +100С. Побеги, пробиваясь на поверхность на поверхность почвы, образуют розетку листьев.
Глазомерная оценка численного обилия сорных растений показала, что в 2009 г. Было явное преобладание по числу особей Sonchus arvensis, и много рассеянных по площади Avena fatua. Остальные виды сегетального сообщества встречались редко или единично. Все сорные растения в период кущения пшеницы занимали нижний ярус, за исключением Sonchus arvensis и, которые занимали ярус культурного растения, а у отдельных особей уже началась фаза бутонизации.
Видовые особенности костей черепа
Кость
Собака
Свинья
КРС
Лошадь
Затылочная кость
Яремные отростки короткие и прямые, имеется затылочный и наружный сагиттальный гребни. Чешуя треугольной формы. Присутствуют мыщелковые отверстия.
Яремные отростки короткие и загнуты медиально. Тело кости цилиндрическое. Затылочный гребень отсутствует. Присутствуют мыщелковые отверстия.
Яремные отростки прямые.
Тело кости цилиндрическое. имеется затылочный и наружный сагиттальный гребни; затылочный хорошо выражен.
Клиновидная кость
Височные крылья мощнее глазничных. Спинка турецкого седла сильно развита. В основании височного крыла находится овальное и каудальное крыловое отверстие. а между височными крыльями и барабанной частью – сонное отверстие.
Глазничное и круглое отверстия сливаются в единое глазничнокруглое. Овальное отверстие редуцированно в овальную вырезку.
В основании височного крыла находится овальное отверстие. Спинка турецкого седла. Глазничное и круглое отверстия сливаются в единое глазничнокруглое.
В основании крыловидного отростка проходит крыловой канал. Спинка турецкого седла слабо выражена. По каудальному краю височного крыла видны овальная и сонная вырезки.
Межтеменная кость
Снаружи сердцевидной формы. На мозговой поверхности выступает кожный мозжечковый намет.
Отсутствует.
Треугольной формы.
Четырехугольная. На мозговой поверхности выступает кожный мозжечковый намет.
Теменная кость
Разделена на две половины сагиттальным гребнем.
У взрослых животных имеет пазуху, которая сообщается с лобной и затылочной пазухами.
Лежит в затылочной и височных областях, разделенная височным гребнями.
На кости виден наружный сагиттальный гребень, который рострально переходит в наружный лобный гребень
Кость
Собака
Свинья
КРС
Лошадь
Височная кость
Чешуя и каменистая часть срослись. Барабанный пузырь круглый без мышечного и подъязычного отростков. Наружный слуховой проход обширный. Скалистый гребень хорошо выражен.
Чешуя и каменистая часть сросшиеся. Височный ход и мышечный отросток отсутствуют. Наружный слуховой проход тонкий и длинный. Барабанный пузырь вытянут ростровентрально.
Чешуя и каменистая часть сросшиеся. Наружный слуховой проход длинный, направлен латерально. Рваное отверстие щелевидное. Барабанный пузырь большой, сплющен; на нем выделяется подъязычный отросток.
Наружный слуховой проход короткий, широкий. Чешуя и каменистая часть срастаются лишь к старости. Рваное отверстие обширное.
Лобная кость
Скуловой отросток не доходит до скуловой дуги. Надглазничное отверстие отсутствует.
Скуловой отросток не доходит до скуловой дуги. От надглазничного отверстия желоб идет только вперед. Лобные пазухи обширные, соединяются с пазухами теменных и затылочных костей.
Очень хорошо развиты. Имеется задний лобный гребень; по его бокам располагаются роговые отростки. Надглазничное отверстие находится в плоскости скулового отростка, рострально и каудально от него проходит желоб, а по направлению к глазнице – канал.
Скуловой отросток лобной кости соединяется с лобным отростком височной кости. Лобные пазухи хорошо выражены.
Носовая кость
Ростральный конец расширен и мелкой вырезкой разделен на два отростка, из которых более длинный латеральный. Носочелестная вырезка отсутствует.
Узкая, длинная, равномерной ширины; на ее наружной поверхности проходит надглазничный желоб. Ростральный конец сужен.
Ростральный конец раздвоен. На внутренней поверхности выступает раковинный гребень.
Ростральный конец заострен, каудальный – огруглорасширенный. На вогнутой внутренней поверхности возвышается дорсальный раковинный гребень.
Сошник
Не прикрепляется к небным костям.
Очень длинный и достигает резцовых костей.
Не прикрепляется к небным костям.
Соединяется с костным небом, начиная от края хоаны.
Кость
Собака
Свинья
КРС
Лошадь
Слезная кость
Не имеют лицевой части, не доходят до носовых костей, но граничат с небными костями. Слезное отверстие широкое.
Лицевая часть участвует в образовании крыловой ямки. Отверстие слезного канала двойное и лежит на глазничном крае. На лицевую часть продолжается крыловая ямка верхней челюсти; каудально от ямки выступает слезный бугорок.
Лицевая часть длинная. У взрослых животных глазничная часть имеет тонкостенный слезный пузырь, который сообщается с челюстной пазухой.
Лицевая и глазничная части одинаковы. Рядом с ямкой слезного мешка имеется плоская мышечная ямка. Содержат ростральные и каудальные слезные бугорки. Отверстие слезного канала воронкообразное.
Верхняя челюсть
Верхнечелюстной бугор небольшой, подглазничный канал короткий, открывается на уровне третьего коренного зуба. Челюстная пазуха слабо развита. Большое нёбное отверстие находится на границе с нёбной костью.
Челюстной бугор мощный, сжатый с боков. Луночек для коренных зубов семь. В нёбном отростке имеется большое нёбное отверстие и нёбный желоб. На лицевой поверхности носовой пластинки видна клыковая ямка, простирающаяся на слёзную и скуловую кости, челюстная пазуха небольшая.
Относительно короткая. На уровне первого коренного зуба расположено подглазничное отверстие, а каудально от него, на уровне третьего коренного зуба — лицевой бугор. Челюстная пазуха обширная, каудально переходит в пазуху слёзного пузыря. В толще нёбного отростка находится нёбная пазуха, сообщающаяся с пазухой нёбной кости. Ростральное нёбное отверстие открывается на нёбной кости. Имеется межальвеолярный край. Челюстной бугор слабо развит, но снабжён особым крыловидным отростком.
Массивная. Мощный лицевой гребень переходит на скуловую кость. Имеется беззубый край, впереди него на границе с резцовой костью у самцов имеется луночка для клыка. Челюстной бугор хорошо выражен. Челюстная пазуха обширная, простирается в челюстной бугор, слёзную и скуловую кости. На нёбном отростке заметен нёбный желоб, который следует рострально от нёбного отверстия.
Кость
Собака
Свинья
КРС
Лошадь
Резцовая кость
Тело кости уплощённое с четырьмя луночками (три резца + клык). Носовой отросток длинный, клинообразный. Нёбный отросток сжат с боков, короткий. Нёбная щель небольшая.
Тело кости массивное, сжато с боков; с тремя зубными луночками для резцов, разрежённо поставленными одна от другой. Нёбные отростки сильно сжаты с боков. Носовой отросток массивный, широкий.
Тело кости пластинчатое; зубные луночки отсутствуют. Между телами правой и левой костей остаётся широкая резцовая щель.
Тело кости массивное с тремя луночками для резцов и половиной луночки для клыка (жеребцы и дикие породы лошадей); каудально от зубных луночек находится беззубый край. Между телами правой и левой костей проходит резцовый канал.
Небная кость
Большая горизонтальная пластинка с несколькими отверстиями в небный канал.
Горизонтальная пластинка мощная, а вертикальная слабая, но содержит пазуху.
Сильно развита. В горизонтальной пластинке заключена пазуха.
Горизонтальная пластинка простирается в виде узкой ленты. Вертикальная — мощная, имеет пазуху.
Скуловая кость
Изогнута в латеральную сторону. Ростральный конец её отдаёт два отростка — слёзный и челюстной; у каудального конца также имеются лобный и височный отростки.
Лицевая часть хорошо выражена и принимает участие в формировании крыловой ямки. Глазничный край снабжён небольшим лобным отростком. Височный отросток сильно развит.
Имеет два каудальных отростка: из них лобный соединяется со скуловым отростком лобной кости, а височный отросток — со скуловым височной кости. Лицевая часть кости развита; на ней имеется скуловой гребень и поверхность для прикрепления большой жевательной мышцы.
Лицевая часть кости сильно развита; на всём протяжении ясно выражен скуловой гребень, переходящий в лицевой гребень верхнечелюстной кости.
Кость
Собака
Свинья
КРС
Лошадь
Нижняя челюсть
Вентральные края несколько выпуклые. На ветви выражен угловой отросток. Ямка для жевательной мышцы глубокая. Мышечный отросток широкий, высокий и слегка отогнут назад. Луночек для коренных зубов семь.
Нижний край тела сильно изогнут вентрально, и несёт пологую сосудистую вырезку. Мышечный отросток длинный, конец его заострён и загнут назад. Суставной отросток седлообразный. Луночек для коренных зубов шесть.
Впереди челюстного утла, на уровне шестого коренного зуба, видна сосудистая вырезка. Мышечный отросток пластинчатый, узкий, длинный, с закругленным концом. Луночка для клыка имеется у самца.
Подъязычная кость
Прикрепляется к подъязычному отростку височной кости. Большие и малые рога цилиндрические. Дистальный членик почти равен среднему, оба имеют вид вытянутых косточек.
Длинным проксимальным члеником прикрепляется к подъязычному отростку височной кости. Тело кости массивное, малые рога соединены с телом хрящами. Дистальный членик ветви представлен короткой эластической связкой. Средний членик тонкий, столбикообразный.
Прикрепляется к шиловидному отростку височной кости. Спереди тела кости выступает язычный отросток. Дистальный членик и малые рога почти равны по длине. Средний членик длинный, пластинчатый, на его свободном конце хорошо выражен мышечный отросток.
Прикрепляется на шиловидных отростках, тело подъязычной кости снабжено спереди длинным язычным отростком. Дистальный членик ветви очень короткий. Средний членик длинный, пластинчатый, узкий, задний конец с мышечным отростком.
Словарная работа в детском саду Словарная работа в детском саду — это планомерное расширение активного словаря детей за счет незнакомых или трудных слов, которое идет одновременно с ознакомлением с окружающей действительностью, воспитанием правильного отношения к окружающему...
Правила наложения мягкой бинтовой повязки 1. Во время наложения повязки больному (раненому) следует придать удобное положение: он должен удобно сидеть или лежать...
Характерные черты немецкой классической философии 1. Особое понимание роли философии в истории человечества, в развитии мировой культуры. Классические немецкие философы полагали, что философия призвана быть критической совестью культуры, «душой» культуры.
2. Исследовались не только человеческая...
Обзор компонентов Multisim Компоненты – это основа любой схемы, это все элементы, из которых она состоит...

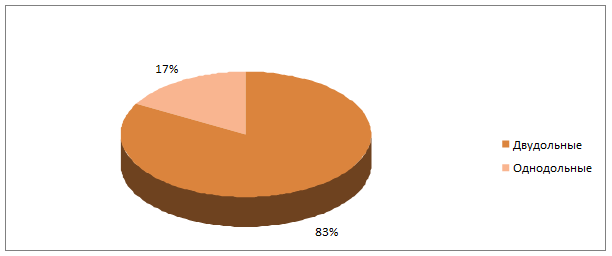 Рис. 1.Соотношение ботанических классов сорных растений в пшеничном агрофитоценозе за годы исследований.
Рис. 1.Соотношение ботанических классов сорных растений в пшеничном агрофитоценозе за годы исследований.