ЭПИДЕРМИС
Функции нервной системы — восприятие внешних раздражений и передача возникающих возбуждений к клеткам, органам, тканям, а также объединение и согласование деятельности отдельных систем органов и организма в целом в единую функционирующую живую систему. Эмбрионально нервная система позвоночных возникает, так же как и у бесчерепных, в виде закладывающейся в эктодерме на спинной стороне зародыша полой трубки. В последующем происходит её дифференцировка, приводящая к образованию: а) центральной нервной системы, представленной головным и спинным мозгом, б) периферической нервной системы, состоящей из нервов, отходящих от головного и спинного мозга, и в) симпатической нервной системы, состоящей в основе из нервных узлов, расположенных около позвоночного столба и связанных продольными тяжами. Головной мозг представлен у позвоночных животных пятью отделами: передним, промежуточным, средним, мозжечком и продолговатым мозгом. Он закладывается эмбрионально в виде вздутия переднего отдела нервной трубки, которое вскоре делится на три первичных мозговых пузыря
ПОКРОВНЫЕ ТКАНИ Покровные ткани расположены на границе с внешней средой, т. е. на поверхности всех органов растения. Они защищают внутренние структуры растений от неблагоприятных внешних воздействий - излишнего испарения и иссушения, проникновения микроорганизмов, резкой смены температуры и т. д. В то же время покровные ткани осуществляют связь растения с окружающей средой. Эти на первый взгляд прямо противоположные функции обусловливаются в каждом конкретном случае специфической структурой клеток. Покровные ткани принимают также участие в метаболических процессах. ОБЩАЯ ХАРАКТЕРИСТИКА, КЛАССИФИКАЦИЯ В связи с выполняемыми функциями строение покровных тканей имеет следующие характерные особенности: а) клетки их соединены плотно, без межклетников; б) клеточные оболочки (чаще всего наружные) обычно утолщаются и претерпевают различные видоизменения, инкру-стируясь суберином, лигнином, кутином, минеральными солями, что повышает их защитные свойства; в) для сообщения с внешней средой образуются специальные структурно оформленные отверстия в виде устьиц или чечевичек; г) покровные ткани относятся к многофункциональным, одна и та же ткань может выполнять функции газообмена, всасывания, защиты от перегрева, выделения и т. д. Поэтому классификация их ведется по преобладающей функции; д) в процессе онтогенеза покровные ткани закономерно сменяют одна другую или одна и та же ткань меняет свою функцию с возрастом. В зависимости от происхождения из различных меристем (генезиса) и строения выделяют три характерных, структурно отличающихся типа покровных тканей - эпидермис, перидерму и корку. Эпидермис - первичная покровная ткань, развивается на листьях и молодых стеблях; перидерма и корка - вторичные покровные ткани - последовательно образуются на стеблях и корнях с возрастом. В некоторых случаях выделяют группу так называемых пограничных тканей, куда относят собственно покровные, или наружные, ткани (эпидермис, перидерма, корка) с преобладающей функцией газообмена и транспирации; наружные ткани с преобладающей функцией всасывания (ризодермис, покрывающий молодые корешки, веламен); внутренние ткани с преобладающей функцией регуляции прохождения веществ (эндодерма, экзодерма в молодых корешках, обкладочные клетки в проводящих пучках листьев) (А. Е. Васильев, Н. С. Воронин, А. Г. Еленевский, Т. И. Серебрякова, 1978). Вся группа пограничных тканей в данном случае рассматривается как физиологический барьер, регулирующий скорость и избирательность проникновения веществ через них. Поскольку такая классификация научно обоснована, но несколько условна и громоздка, авторы впоследствии отказались от нее. В настоящем учебнике ризодермис, веламен, эндодерма, экзодерма рассматриваются при изучении соответствующих органов. ЭПИДЕРМИС Эпидермис (эпидерма, кожица; греч. epi - над, сверху и derma - кожа) - самый наружный слой клеток растений, образующийся из прото-дермы конуса нарастания и имеющий первичное строение. В редких случаях эпидермис может иметь вторичное строение. Так как тело растения - открытая живая система, непрерывно взаимодействующая с окружающим миром, в процессе эволюции возникла особая ткань, лежащая на границе окружающей, среды и внутренней структуры растения и регулирующая их взаимосвязь. Без эпидермиса существование высших наземных растений было бы невозможно. Первые структурно довольно примитивные сухопутные растения - ринии - обладали вполне типичным эпидермисом. Особая функция эпидермальной ткани, расположенной на границе двух сред, осуществляется благодаря специфической структурной организации. Плотно расположенные клетки эпидермиса образуют покров, защищающий растение от механических повреждений, иссушения, проникновения микробов, грибов-паразитов и т. д. Другие структурные особенности кожицы - кутикула, воск, волоски - усиливают защитные функции. В то же время живые, функционирующие клетки эпидермиса, изолируя внутренние ткани, обладают повышенной проницаемостью для целого ряда веществ, в том числе для углекислого газа, кислорода, паров воды, интенсивный обмен которых происходит в процессе фотосинтеза, дыхания и транспирации (лат. trans - через и spiio - дышу, выдыхаю; физиологическое испарение воды растением). Это противоречие разрешается с помощью высокоспециализированных устьичных клеток, между которыми находится отверстие - устьичная щель. Щели периодически открываются и закрываются, регулируя газообмен и обмен паров воды. Защита растения от неблагоприятных внешних факторов и регуляция газо- и парообмена - важнейшие функции эпидермиса. Кроме того, ткань кожицы может выделять наружу различные вещества (соли, воду, эфирные масла), принимать участие в фотосинтезе, поглощении воды и питательных веществ, синтезе различных соединений, в движении листьев, воспринимать раздражение и т. д.
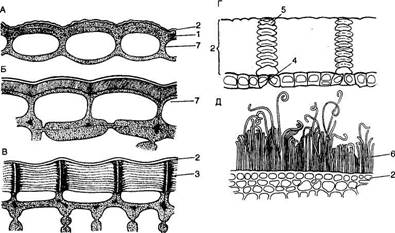
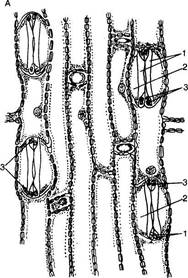
Полифункциональность эпидермиса обусловливает его строение. Это сложная ткань, так как состоит из морфологически разнородных элементов: 1) основные эпидермальные клетки; 2) устьичный комплекс; 3) выросты эпидермиса в виде различного типа волосков. Кроме того, в состав эпидермиса некоторых растений входят моторные клетки, парные клетки и окремневшие бугорки. Основные эпндермальные клетки. Являются относительно неспециализированными и слагают массу кожицы. Чаще всего они имеют таблитчатую форму и всегда живое содержимое. Снаружи очертания эпидермальных клеток в разных органах и у разных растений чрезвычайно разнообразны. Размеры и очертания клеток кожицы формируются в большой зависимости от соотношения скорости роста органа в целом и его поверхности. Поэтому в удлиненных частях рас-тзния (стебли, черешки, жилки листа, листья большинства однодольных) эпидермальные клетки вытянуты в направлении длинной оси органа (рис. 71, А, Б). В листьях, длина которых равна или ненамного превышает ширину, а также в лепестках, завязях, семяпочках эпидермальные клетки часто имеют волнистые боковые стенки (антиклинальные, т. е. направленные перпендикулярно поверхности органа), что повышает прочность эпидермиса (рис. 71, В, I). Степень волнистости стенок часто зависит от условий произрастания растений. У растений, выросших при интенсивном освещении, волнистость выражена слабфе по сравнению с затененными. Все гипотезы, объясняющие причины волнистости, пока мало обоснованны. В литературе имеются попытки классифицировать эпидермис по степени извилистости клеток - прямолинейный, криволинейный и разнолинейный (по Н. А. Анели, 1970). Органы одного и того же растения покрыты эпидермальными клетками разных формы и размеров (рис. 72). Наиболее мелкие клетки характерны для растений аридных (лат. aridus - сухой) районов (степи, пустыни). Основные клетки эпидермиса прочно соединены друг с другом и, как правило, не имеют межклетников. Рядом специфических особенностей обладают оболочки эпидермальных клеток. Наиболее сложную организацию имеют внешние стенки. У водных растений и растений, оптимальных условий местообитаний (мезофиты) эпидермис в целом тонкостенный, наиболее утолщенными являются наружные стенки оболочки. Эпидермис с чрезвычайно утолщенными оболочками свойствен листьям хвойных. Важнейшая функция эпидермиса — ограничение транспирации — в большей степени обусловливается наличием жирового вещества кутина. Это вещество инкрустирует наружную стенку или образует самостоятельный слой — кутикулу — на поверхности эпидермиса. В процессе кутшизации происходит пропитывание целлюлозной оболочки кутаном. Комплекс кутикулы и кутинизированной оболочки имеет сложную организацию. В его состав входят целлюлоза, пектиновые соединения, кутан, воск и некоторые другие инкрустирующие вещества.
У тропических растений, например, слой кутикулы и воска достигает 0,2— 0,5 см. Эпидермис листьев и стеблей водных растений почти не имеет кутикулы и тем более воскового налета. Поверхность кутикулы может быть гладкой или на ней образуются разнообразные выступы, гребни, трещины, складки. Воск чаще всего образует тонкий мелкозернистый налет либо чешуйки, палочки и другие структуры различных очертаний (рис. 74). Важной особенностью кутикулы является то, что во влажном состоянии она более проницаема для газов и жидкостей, а при подсушивании проницаемость резко снижается. Таким образом, комплекс кутикулы и кутинизированной оболочки представляет покров, защищающий растение от иссушения. Кроме того, он предохраняет от заражения всевозможными грибами-паразитами, бактериями, вирусами, которые в изобилии находятся на поверхности растения. Обладая водоотталкивающими свойствами, инкрустированная кутином и воском оболочка плохо смачивается, и попавшие на поверхность растения капли воды легко с нее скатываются. В связи с этим развитие и проникновение внутрь болезнетворных организмов становится затруднительным.
В то же время в некоторых местах наружные стенки эпидермиса обладают повышенной проницаемостью, обусловленной характером распределения в них кутина и воска, числом и характером пор. Оболочка клеток эпидермиса, особенно наружная, может пропитываться солями кальция или соединениями кремния (хвощи, осоки, злаки). У некоторых растений оболочки эпидер-мальных клеток надземных органов одревесневают, утолщаются, что сокращает размеры клеточных полостей. При этом часто одревесневают и клетки эпидермиса, и клетки подстилающего его слоя. Строение оболочек эпидермальных клеток непрерывно меняется с возрастом и под влиянием условий жизни. Эти изменения обусловлены деятельностью живого протопласта эпидермиса, имеющего довольно сложную организацию. Нормально развитая клетка эпидермальной ткани заполнена крупной вакуолью с клеточным соком, бесцветным или окрашенным пигментом. Протопласт занимает пристенное положение, ядра чаще всего располагаются около внутренних стенок. Пластидный аппарат у большинства растений представлен хлоропластами, которые имеют небольшое количество слаборазвитых гран. У светолюбивых растений пластиды часто разрушаются, сохраняясь иногда в клетках нижнего эпидермиса. У растений, произрастающих в тени, хлоро-пласты могут присутствовать во всех эпидермальных клетках. У некоторых растений пластиды в эпидермисе листьев сохраняются в виде мелких шаровидных лейкопластов, в периферической части пластид обнаружены трубочки, сходные по внешнему виду с микротрубочками. Полагают, что такие трубочки принимают участие в транспорте белков, синтезируемых пластидой. Лейкопласты эпидермальных клеток часто содержат крупные крахмальные зерна, митохондрии обладают хорошо развитой системой крист. Аппарат Гольджи активный, от цистерн диктиосом отчленяется большое число крупных пузырьков. У многих растений обнаружен хорошо развитый гранулярный эндоплазматический ретикулум. Эти особенности структуры протопласта свойственны секреторным клеткам, следовательно, основные эпидермальные клетки обладают чертами строения, характерными для железистых тканей. Это дает основание охарактеризовать основные клетки эпидермиса как метаболически активные, принимающие участие в биосинтезе и выделении определенных веществ (кристаллы солей, ароматические вещества и т. д.). В некоторых случаях эпидермис состоит из нескольких рядов клеток (от 2 до 15-16). При образовании многослойного эпидермиса протодерма делится несколько раз параллельно поверхности листа. Предполагают, что основная функция такого типа кожицы — запас воды, поэтому он встречается преимущественно у тропических растений, обитающих в условиях непостоянной обеспеченности водой (пальмы, орхидеи, фикус, пеперомия и др.). От многослойного эпидермиса надо отличать гиподерму (греч. hypo — под, внизу и дерма) — ткань с малым количеством хлоропластов или совсем без них, расположенную непосредственно под эпидермисом листьев, хвои, семян (рис. 75). Гиподерма возникает независимо от эпидермиса, отличается по строению и от клеток кожицы, и от глубжележащих тканей. Функции различные: часто ее клетки с сильно утолщенными оболочками усиливают механические и изолирующие свойства эпидермиса. В ней могут накапливаться пигменты, таннины, алкалоиды и т. д.
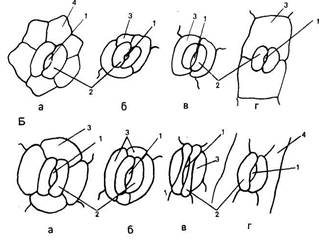
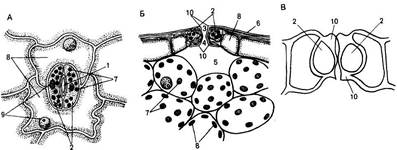
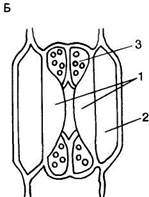
Устьица. Одна из основных функций эпидермиса, регуляция газообмена растений, осуществляется с помощью системы высокоспециализированных клеток — устьиц, резко отличающихся по структуре от основных эпидермальных клеток. Устьица представляют собой отверстия (устьичные щели, или апертуры), ограниченные двумя специализированными клетками эпидермиса, которые называют замыкающими. Термин «устьице» обозначает весь комплекс - отверстие и замыкающие клетки. Обычно устьица окружены клетками, не отличающимися от других эпидермальных клеток. У многих растений, наоборот, к устьицам примыкают отличающиеся по форме и иногда по содержимому клетки эпидермиса. Эти особые клетки носят название сопровождающих или побочных. Они участвуют в изменении осмотического давления, регулирующего изменение формы и движение замыкающих клеток, которые открывают или закрывают устьичную щель. Форма и число клеток устьичного комплекса могут быть различными, на основании этого различают типы устьиц. (У двудольных растений чаще всего используется классификация устьиц, предложенная в 1950 г. К. Меткафом и Л. Чоком в «Анатомии двудольных». Они выделяют четыре основных типа устьиц (рис. 76, А). 1. Аномоцитный тип - устьица окружены неопределенным числом клеток, не отличающихся по форме и размерам от остальных эпидермальных клеток (76, А, а). 2. Анизоцитный тип - имеются три околоустьичные клетки, при этом одна из них гораздо меньше двух других (76, А, б). 3. Парацшпный тип - сопровождающие клетки располагаются вдоль замыкающих клеток (76, А, в). 4. Диацитный тип - замыкающие клетки окружены двумя побочными, смежные стенки которых располагаются под прямым углом к длинной оси устьица (76, А, г). У однодольных различают следующие типы устьиц, выделенные Г. Стеббинсом и Г. Кушем (1961) (рис. 76, Б). 1. Замыкающие клетки окружены четырьмя или шестью сопровождающими, которые образуют кольцо (76, Б, а). 2. Замыкающие клетки окружены четырьмя или шестью побочными, две из которых значительно меньше других и располагаются у концов замыкающих клеток. Остальные располагаются параллельно длинной оси устьичной щели вдоль замыкающих клеток (76, Б, б). 3. 3. Замыкающие клетки сопровождаются двумя побочными, длинные оси которых направлены параллельно длинной оси устьичной щели (76, Б, в). 4. 4. Околоустьичные клетки отсутствуют (76, Б, г). Существуют и другие классификации устьиц. Так, согласно новой морфологической классификации Дж. Пэтела (1979), выделяется 44 типа устьиц, отличающихся числом, размерами, положением и формой побочных клеток. У двудольных наиболее распространен парацитный тип устьиц (рис. 77). Замыкающие клетки имеют почковидную (бобовидную) форму - такими они видны с поверхности листа (рис. 77, А). Наружные стенки замыкающих клеток обычно имеют выросты, что хорошо видно на поперечном срезе устьица (рис. 77, Б). Пространство, ограниченное этими выростами, называют передним двориком. Нередко аналогичные выросты наблюю
даются и у внутренних оболочек замыкающих клеток. Они образуют задний дворик, соединенный с крупным межклетником - подустъичной полостью. Движение устьиц обусловливается изменением тургорного давления в замыкающих клетках (см. дальше), однако способы их открывания и закрывания зависят от строения. У большинства растений оболочка замыкающих клеток неравномерно утолщена. Рассматриваемый устьичный комплекс по способу открывания относится к типу амариллисовых. Спинные стенки замыкающих клеток очень тонкие, брюшные имеют утолщения, образованные в основном верхними и нижними выростами (рис. 77, В). При повышении тургорного давления тонкие спинные стенки сильно растягиваются, вдавливаясь в соседнюю клетку. Вследствие этого изгибается и брюшная, более жесткая стенка и устьичная щель открывается. Наружная стенка клеток покрыта кутикулой, нередко распространяющейся и на поверхность, обращенную к устьичной щели и подустьичной камере. Устьица могут быть полностью покрыты воском. Каждая из замыкающих клеток имеет крупное ядро и хлоропласты, которые периодически накапливают крахмал. У однодольных наиболее характерным является устьице злаковых (см. рис. 76, Б, в).
Например, считают, что радиальное расположение микрофибрилл целлюлозы в оболочке замыкающих клеток играет более важную роль в движении устьиц, чем неравномерное утолщение оболочки. С помощью поляризационной оптики и электронной микроскопии установлено, что микрофибриллы целлюлозы располагаются в замыкающих клетках как бы по радиусам с центром схождения в области устьичной щели, тогда как в сопровождающих клетках микрофибриллы ориентированы вдоль длинной оси.
Полагают, что устьице открывается при увеличении количества ионов калия, которые активно (т. е. против градиента концентрации и с затратой энергии) поступают в замыкающие клетки из соседних клеток. Это ведет к повышению осмотического давления в клетках и увеличению их объема за счет всасывания воды. Отток ионов происходит пассивно по градиенту концентрации. При этом вода уходит из клеток, объем их уменьшается, устьица закрываются. В замыкающих клетках находятся многочисленные активно действующие митохондрии, поставляющие энергию для транспорта ионов. Высокоспециализированные хлоропласты синтезируют углеводы, необходимые для активной работы митохондрий. Первоначальная гипотеза, до сих пор излагаемая во многих учебниках, о том, что расщепление крахмала, синтезируемого хлоропластами замыкающих клеток, вызывает увеличение осмотического давления вследствие образования сахара, поглощающего воду, в настоящее время сменилась предположением, согласно которому гидролиз крахмала приводит к накоплению органических анионов, с чем и связано поглощение калия. Несомненно, движение устьичного комплекса имеет более сложный характер, зависит от суммы внутренних и внешних (температура, вода) факторов. Детально этот вопрос изучается в курсе физиологии растений. Устьица встречаются на всех надземных частях растений, но преимущественно в эпидермисе листьев. Причем большая их часть сосредоточена на нижней поверхности листьев (гипостомати-ческие листья), что способствует менее интенсивному испарению. Есть и такие растения, у которых устьица образуются лишь на верхней стороне (на листьях, плавающих на поверхности воды, - эпистоматические листья), либо на обеих сторонах (амфистоматические листья). Количество устьиц сильно варьирует в зависимости от вида растения и от условий обитания. У обычных растений леса, луга, у многих культурных растений число их колеблется от 90 до 700 на 1 мм2 поверхности листа, у некоторых насчитывается до 1300 устьиц на 1 мм2. У экземпляров одного и того же вида растений плотность и размеры устьиц в значительной степени зависят от экологических условий. Например, на листьях подсолнечника, растущего в условиях полного освещения, на 1 мм2 листа приходится около 220 устьиц, а при небольшом затенении - около 140. Плотность устьиц обычно возрастает от нижних листьев к верхним. Устьица также различаются по уровню расположения относительно поверхности эпидермиса. Некоторые из них расположены вровень с другими эпидермальными клетками, другие подняты выше или погружены ниже поверхности. У однодольных, листья которых растут преимущественно в длину, устьица образуют правильные параллельные ряды, тогда как у двудольных они разбросаны в различных направлениях, поскольку листья интенсивнее растут в ширину или в плоскостном направлении (см. рис. 77, 78). Число и размеры устьиц реагируют на каждый конкретный комплекс факторов окружающей среды, что дает некоторые представления о характере взаимоотношения каждого растения со средой обитания. Поэтому изучение количества и размеров устьиц (так же, как и других анатомических элементов) относится к категории количественно-анатомических методов, которые используют при экологических исследованиях, а также для характеристики сортов культурных растений. У некоторых растений засушливых местообитаний (ксерофитов) наряду с обычными устьицами встречаются сложные устьичные комплексы. Иногда попадаются и двухэтажные устьица, впервые описанные В. X. Тутаюк, с двумя парами замыкающих клеток и 4-6 сопровождающими клетками, расположенными в два этажа (рис. 80). В. X. Тутаюк наблюдала их у дрока ситниковид-ного, у видов можжевельника, тисса, сосен, у растений глинистых, песчаных и крайне сухих почв во многих районах Азербайджана.
Наряду с устьицами, предназначенными для газообмена и транспирации, у многих растений имеются водные устьица - гидатоды, выделяющие воду в капельно-жидком состоянии (см. Выделительные ткани). Трихомы (греч. trichoma - волосы, волосяной покров). Поверхность эпидермиса надземных органов редко бывает гладкой и однородной, а часто образует разнообразные выросты. Такие выросты встречаются на всех частях растения и могут существовать в течение всей жизни органа или быть эфемерными. Выросты могут образовываться клетками самого эпидермиса трихомы либо клетками более глубоко расположенных тканей - эмергенцы (лат. emergere - выдаваться). Чаще всего все выросты эпидермиса называют трихомами. Они отличаются громадным разнообразием, в связи с чем нередко предпринимались попытки классифицировать их. Наиболее полными являются классификации X. Солередера (1908) и Дж. Упхофа (1962). Все типы трихом делятся на кроющие, не обнаруживающие секреторной активности, и железистые, выделяющие секрет. Морфологически они могут быть представлены различными типами (рис. 81). Одним из наиболее распространенных типов является волосок. По внешнему виду волоски подразделяются на одноклеточные и многоклеточные; одноклеточные могут быть простыми или ветвистыми; многоклеточные - состоять из одного или нескольких рядов клеток. Одни многоклеточные волоски ветвятся по типу деревьев, другие - звездчатые волоски - располагают ветви в одной плоскости. У многоклеточных волосков обычно наблюдаются ножка, погруженная в эпидермис, и тело, выступающее над его поверхностью. Клетки, окружающие ножку, часто отличаются по структуре от остальных эпидермальных клеток и образуют основу («подошву») волоска. Другим распространенным типом трихом является чешуйка, или щитовидный, пельтатный (лат. peltatus - дисковидный или щитовидный), волосок. Чешуйка образована дисковидной многоклеточной пластинкой, сидящей на стебельке или прикрепленной к эпидермису непосредственно основанием. Одноклеточные, многоклеточные и щитовидные волоски могут быть кроющими и железистыми. У кроющих волосков протопласт быстро отмирает и оболочка мертвой клетки заполняется воздухом. Такие блестящие белые волоски покрывают органы многих растений (листья мать-и-мачехи, лоха серебристого, коровяка; черешки листьев; стебли гераней, лапчаток и т. д.), образуя шерстистый, войлочный или иной покров. У злаков, многих зонтичных, бобовых кроющие волоски остаются жизнедеятельными в течение продолжительного времени. Протопласт таких волосков отличается своеобразным строением: крупные ядра, мелкие пластиды с плотным матриксом и единичными тилакоидами, много-
численные митохондрии с хорошо развитой системой крист. Основной объем цитоплазмы занимает агранулярный эндоплазматический ретикулум. Комплекс Гольджи развит слабо, липидные капли достигают больших размеров. Анализируя структуру протопластов кроющих волосков у многих растений, ряд авторов (А. Е. Васильев, Ю. В. Гамалей, М. Ф. Данилова, Г. М. Козубов, 1980) приходят к заключению, что на ранних стадиях развития кроющие волоски функционируют как метаболически активные образования, а после отмирания протопластов - как защитные. Высказывается предположение, что между живыми кроющими и железистыми волосками не существует резких функциональных различий. Железистые волоски обычно сохраняют живое содержимое клеток. Некоторые из простых многоклеточных железистых волосков состоят из ножки и одноклеточной головки, составляющей секреторную часть волоска. У пельтатных железистых трихом дисковидная пластинка состоит из железистых клеток. Выделения железистых волосков разнообразны — смолистые, сахаристые вещества, эфирные масла, слизи, вода с растворенными веществами различной природы. Хорошо известны смолистые вещества, выделяемые железистыми волосками почечных чешуи, которые как бы закупоривают почку и предохраняют нежные листочки от высыхания и вымерзания. Оболочки клеток трихом обычно целлюлозные и покрыты кутикулой. Иногда они одревесневают. Часто у волосков образуется толстая вторичная оболочка. Так, используемые человеком волоски семян хлопчатника представляют собой чрезвычайно длинные эпидермальные выросты с толстыми вторичными оболочками, состоящими почти из чистой целлюлозы. Они образуются из протодермы семяпочки во время цветения, и их развитие продолжается непрерывно в течение 10-12 дней после цветения. В зависимости от сорта хлопчатника волоски достигают в длину 10-70 мм. Вторичную оболочку имеют также цепляющиеся волоски хмеля. Значение волосков разнообразно и во многих случаях еще не выяснено. Принято считать, что наиболее распространенной функцией волосков типа кроющих является предохранение поверхности тела от излишнего испарения. Блестящие белые волоски, с одной стороны, отражают прямые солнечные лучи, а с другой — густым сплетением создают над эпидермисом среду, способствующую уменьшению транспирации. Однако в ряде случаев, у частей цветка, покров из волосков повышает интенсивность транспирации. Е. А. Мирославов (1965) считает, что кроющие волоски способны снижать интенсивность транспирации лищь в тех случаях, когда образуют густой войлочный покров. Жизнедеятельные же волоски, наоборот, повышают интенсивность транспирации, поскольку они во много раз увеличивают поверхность органа. К. Эсау (1980), рассматривая функции большинства типов трихом, выделяет следующие: 1) защита органа от перегрева; 2) выведение солей из ткани листа и в связи с этим предотвращение накопления токсичных солей в растении; 3) механическая и химическая защита от насекомых. В. Г. Александров (1966) описывает разнообразные по структуре и содержимому волоски, которые появляются на цветках злаков и отмирают после оплодотворения, несомненно, оказывая влияние на важнейшие процессы опыления цветков. Чувствительные трихомы, содействующие опылению цветка, отмечены на тычиночных нитях василька, портулака (Г. Габерландт, 1924). По наблюдениям В. Г. Александрова и М. И. Савченко (1947), на семенах некоторых видов ромашек, произрастающих в пустыне, находятся волоски, впитывающие атмосферную влагу, которая выпадает в виде капель росы и дождя. Оболочки клеток этих трихом имеют сильно выраженную складчатость и лишены кутикулы. При доступе воды оболочки трихом ослизняются и, набухая, впитывают влагу. С наступлением сухого периода поверхностные участки клеток подсыхают, образуя непроницаемую для воды пленку, и поглощенная влага просачивается во внутренние ткани. Приведенные примеры дают некоторое представление о функциях трихом, все разнообразие которых недостаточно изучено даже в специальной, научной литературе. Отличаясь чрезвычайным многообразием, трихомы специфичны для определенных таксонов, поэтому часто используются в систематике растений в качестве диагностических признаков. Более детально функции железистых волосков, участвующих в биосинтезе и секреции определенных веществ, и корневых волосков будут рассмотрены в соответствующих разделах книги. Эмергенцы. Представлены на поверхности эпидермиса особыми выростами, в формировании которых, кроме кожицы, принимают участие лежащие под ней клетки. К ним относятся шипы розы, малины, ежевики, покрывающие черешки листьев и молодые побеги. Иногда к эмергенцам относят волоски хмеля, шипы на плодах многих зонтичных, каштана конского, дурмана. Существуют переходные формы между эмергенцами и трихомами. Эмергенцы располагаются на эпидермисе без определенного порядка и этим отличаются от морфологически сходных с ними колючек. Эпидермис образуется не только снаружи органа, но и внутри, если в органе формируется полость. Например, ткань, выстилающая полость плодов (гороха, бобов, фасоли, мака, дурмана и т. д.), по своему происхождению идентична эпидермису наружной стенки плода. Строение внутреннего эпидермиса в связи с особыми условиями (повышенная влажность, отсутствие света и т. д.) довольно существенно отличается. У плодов гороха, например, образуется большое число волосков, расположенных особенно густо около семян и, несомненно, принимающих физиологическое участие в процессе их развития. У мака, многих гвоздик, ирисов, дурмана оболочки внутреннего эпидермиса значительно утолщаются и одревесневают ко времени созревания плодов. Волоски развиты слабо, размеры их невелики, кутикула выражена более заметно, клетки эпидермиса плотно сомкнуты, иногда среди них встречаются устьица. Продолжительность существования эпидермиса на надземных побегах обычно ограничивается одним вегетационным периодом. Однако у растений южных регионов многолетний эпидермис характерен для листьев (лавр, лавровишня, магнолия), побегов (эвкалипт, платан, стеркулия). Многолетний эпидермис на хвое сосны, ели, туи, на стволах черемухи, ольхи серой. На поверхности многолетнего эпидермиса развивается толстая кутикула. У хвойных оболочки эпидермальных клеток одревесневают.
|
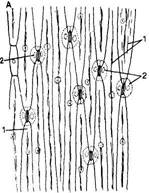

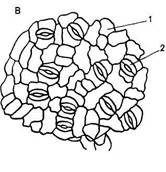
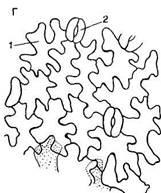
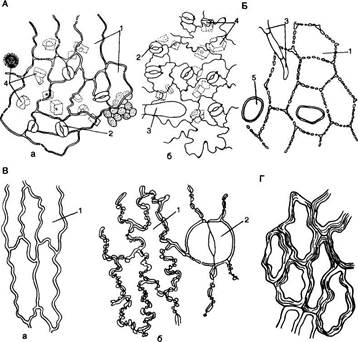
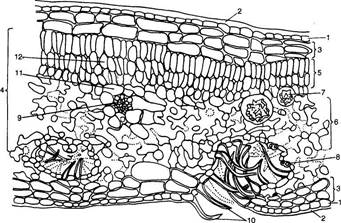
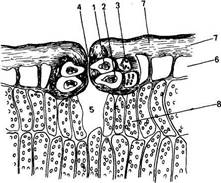
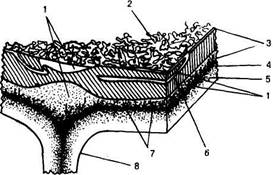 Рис. 73. Схема организации наружной оболочки клеток верхнего эпидермиса листа груши - Purus communis (по R. F. N orris, М. J. Bukovas, 1968):
1 - кутиновый матрнкс; 2 - эпикутнкулярный воск; 3 - кутикула; 4 - пектиновые вещества; 5 - целлюлозная оболочка; 6 - двоякопреломляющий слой воска, погруженный в куши; 7 - пектиновые канальцы; 8 - антиклинальная стенка
Рис. 73. Схема организации наружной оболочки клеток верхнего эпидермиса листа груши - Purus communis (по R. F. N orris, М. J. Bukovas, 1968):
1 - кутиновый матрнкс; 2 - эпикутнкулярный воск; 3 - кутикула; 4 - пектиновые вещества; 5 - целлюлозная оболочка; 6 - двоякопреломляющий слой воска, погруженный в куши; 7 - пектиновые канальцы; 8 - антиклинальная стенка