
Часть 4. Вкус у млекопитающих
Вкус обычно рассматривается как контактное чувство. Действующие молекулы находятся в растворе, который контактирует с рецептором. Напротив, обоняние – дистантное чувство. Молекулы химических веществ доставляются к обонятельным рецепторам с потоками воздуха. Очевидная условность такой классификации – на клеточном, а тем более на молекулярном уровне рецепторный механизм может быть одним и тем же. Вкус хорошо развит у позвоночных. Как и все другие животные, человек получает жизненно важную информацию о пище, уже когда она оказывается во рту – в последний момент перед тем, как сознательный контроль над процессом потребления пищи будет утрачен Не удивительно, что вкус имеет сложную нейрологическую основу. Импульсация идет в основном от вкусовых рецепторов языка, но также от надгортанника, верхней трети пищевода и мягкого нёба. Одновременно поступает важная информация о характере поверхности пищи, текстуре, температуре и запахе; свою роль играют и память, и культура воспитания – то, что вкусно в одной культурной традиции, то отвратительно – в другой. Вкусовые ощущения, как и запахи, многообразны. В этом отношении они отличаются от механочувствительности, такой ее формы, как осязание. Осязательное же ощущение всегда имеет одно и то же качество и различается только по интенсивности. Запахи же различаются и по количественной, и по качественной осям. В этом отношении оба чувства – вкус и обоняние – больше напоминают такую сложную форму механочувствительности, как слух; есть черты сходства и со зрением. Современные воззрения восприятия вкусовых ощущений восходят к идее Ханса Хеннинга, высказанной в 1922 году, что вкусовое ощущение человека может быть разложено на четыре основных: сладкое, соленое, кислое и горькое. В последнее время исследователи выделяют также приятный вкус глютамата натрия, называемый по-японски umami, который существенно отличается от четырех классических вкусов, а также вкус воды. Эта субъективная классификация связана и с объективной биологией. Позитивные ощущения сладкого или umami связаны с высококалорийной легко усвояемой пищей. Фрукты и ягоды часто рекламируют это яркими окрасками, адресованными зрению. Соленость, ощущение кислого и вкус воды связаны с регуляцией ионного состава «внутренней среды», в частности рН. Горечь воспринимается как сигнал тревоги – поедаемый материал опасен. Такой объективный анализ предполагает, что и у других животных присутствует схожая (хотя и далеко не идентичная) организация, что и показано у многих видов позвоночных. Схожи ли их субъективные ощущения, конечно, остается открытым вопросом.
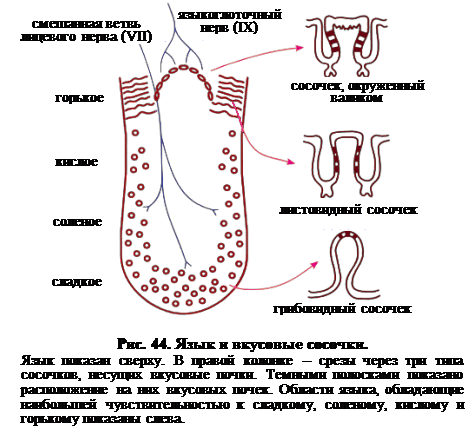
Вкусовые рецепторы группируются во вкусовые почки, локализованные в выступах, именующихся вкусовыми сосочками. На языке человека присутствуют четыре типа сосочков: нитевидные, грибовидные, листовидные и окаймлённые (окруженные валиком) (рис. 44). Нитевидные сосочки никаких вкусовых почек не содержат. Их функция – чисто механическая, они придают языку характер тёрки, что у многих млекопитающих, например у домашней кошки, выражено значительно сильнее, чем у человека. Три других типа сосочков содержат вкусовые почки. Грибовидные сосочки, названные так из-за сходства со шляпкой гриба, группируются главным образом в передней части и на переднем краю языка. Вкусовые почки, чувствительныек сладкому и соленому, погружены в поверхностный эпителий. Вследствие обильного кровоснабжения почки выглядят маленькими красными точками в передней части языка. Листовидные сосочки располагаются в параллельных складках (откуда и название) в количестве 3-8 у корня языка. Эти вкусовые почки особенно чувствительны к кислому. Окаймлённые сосочки, как показывает их название, несколько погружены в поверхность языка и окружены углублением, которое отделяет их от остальной поверхности языка. Восемь-девять таких сосочков образуют V-образную кофигурацию у корня языка. В стенках каждого сосочка и в стенках бороздки, окружающей сосочек, развивается около 250 вкусовых почек. Они наиболее чувствительны к кислому и горькому. В стенках сосочка располагаются также секреторные железы, которые при движениях языка омывают бороздку. Как упоминалось выше, другие вкусовые почки располагаются в надгортаннике, верхней трети пищевода и мягком нёбе.
Иннервация вкусовых почек достаточно сложна: Ø передняя часть языка иннервируется смешанной ветвью лицевого нерва (черепно-мозговой нерв VII), Ø задняя часть языка – языкоглоточным нервом (черепно-мозговой нерв IX). Ø почки, расположенные вне языка, иннервируются ветвью блуждающего нерва (черепно-мозговой нерв X).
Вкусовая почка представляет собой группу из 50-150 клеток, выстроенных подобно долькам апельсина, и окруженную эпителиальными клетками сосочка (рис. 45 В). Вкусовая почка содержит два класса клеток: Ø главные Ø базальные. Главные клетки, в свою очередь подразделяются на три типа: ü тип 1 (темные клетки), ü тип 2 (светлые), ü тип 3. Клетки типов 1 и 3, в основном, являются рецепторными, тогда как клетки типа 2 – обычно опорные. Такое приблизительное описание функций связано с тем, что роль этих клеток варьирует у разных видов животных и даже от одной части языка к другой. Апикальные части всех трех типов главных клеток соединены плотными контактами и выступают во вкусовую пору. Эта пора заполнена секретом и открыта на поверхность сосочка. Клетки типа 1 составляют до 65% клеточной популяции вкусовой почки кролика. Их апикальные части образуют кисточки из 30-40 микроворсинок. В них также содержатся плотные гранулы с секретом, который выделяется во вкусовую пору. Стенки клеток типа 1 имеют многочисленные листовидные выросты, которые охватывают немиелинизированные нервные волокна и клетки типа 2. Синапсы с нейронами клетки типа 1 образуют у мышей, но не у кроликов. Клетки типа 2 составляют прибл. 20% клеточной популяции вкусовой почки. Их апикальные части заполнены электронно-плотным материалом и оканчиваются множеством тупых микроворсинок, направленных во вкусовую пору. Синапсы с нейронами найдены у мышей, но не у кроликов. Клетки типа 3 – менее 10% клеток вкусовой почки у кроликов. У них образуется по одной крупной микроворсинке, направленной во вкусовую пору. В отличие от двух других типов клеток, которые могут образовывать, а могут и не образовывать синапсы, клетки типа 3 всегда образуют синапсы с соседними нейронами и, следовательно, во всех случаях должны рассматриваться как рецепторные клетки.
Все клетки вкусовой почки являются короткоживущими и постоянно замещаются. Радиоактивное мечение эпителиальных клеток, окружающих вкусовую почку, показывает, что дочерние клетки перемещаются во вкусовую почку со скоростью 1 клетка/10 часов. Т.е. средняя продолжительность их жизни во вкусовой почке не превышает 10 дней. За это время клетка перемещается с периферии почки к центру. В конце своей краткой жизни клетки вкусовых почек либо подвергаются фагоцитозу, либо претерпевают апоптоз. Такой быстрый обмен клеток вкусовых почек, вероятно, и объясняет трудность определения точной функции каждого их типа. Вкусовые рецепторные клетки (ВРК) в отличие от обонятельных, являются истинными рецепторными клетками. Волокна сенсорных нервов входят в базальную пластинку почки (где и утрачивают миелиновую оболочку) и достигают клеток почки. Волокна покрыты мембранными выростами клеток типа 1 (темных). Одно волокно обычно иннервирует больше, чем одну вкусовую рецепторную клетку, и, напротив, одна рецепторная клетка контактирует с более, чем одним нервным волокном. Поскольку происходит постоянный обмен клеток вкусовых почек, окончания нервных волокон находятся в процессе постоянного поиска новых синаптических контактов.
До настоящего времени вопрос о том, насколько специализированы вкусовые рецепторные клетки остается дискуссионным. Вопрос о том, отвечает ли клетка только на один или на несколько из первичных шести вкусовых стимулов пока не решен. Общее мнение склоняется в пользу признания генерализованности. Отведения от одиночного нервного волокна из передней части языка крысы или хомячка позволяют идентифицировать некоторые волокна как максимально чувствительные к NaCl, т.е. соленому (N-волокна), с менее выраженным ответом на НС1, т.е. кислое. Другие волокна (Н-волокна) лучше всего реагируют на НС1, более слабо – на NaCl и хинин (горечь). Однако, опять-таки здесь есть масса видовых различий, к тому же следует помнить, что веточки одного и того же волокна могут иннервировать несколько вкусовых рецепторных клеток. Исследование молекулярной биологии и биофизики одиночной вкусовой рецепторной клетки технически трудно, и к настоящему моменту часто дает противоречивые результаты.
Umami – это название вкуса некоторых видов японской пищи – сушеной рыбы кацуво (Katsuwonuspelamis), морской капусты комбу (Laminariajaponica) и т.д. Этот вкус присущ также и куриному бульону. Вкус этот существенно отличается от сладкого, хотя тоже воспринимается как приятный. Наиболее значительным активным ингредиентом, имеющим этот вкус, является l-глютамат, хотя некоторые другие аминокислоты, короткие пептиды и нуклеотиды производят аналогичный эффект. Есть данные, что рецепторной молекулой для вкуса umami является глутаматный рецептор, который, возможно, является вариантом глутаматного рецептора ЦНС. Соленость. Детектирование соли для животных имеет большое значение, поскольку является ключевым для ионного гомеостаза внутренней среды. Показано, что Na+ воздействует на чувствительные к соленому вкусовые рецепторные клетки, входя в Na+-специфические ионные каналы (рис. 47), а не включая систему G-белков. Это было продемонстрировано с помощью блокатора Nа+-каналов – амилорида. Это соединение гуанидина является хорошо известным внешним блокатором эпителиальных Na+-каналов (ЭNaK). Эти каналы никак не связаны с потенциал-зависимыми Na+-каналами возбудимой мембраны. В присутствии субмикромолярных концентраций амилорида на слизистой поверхности грибовидного сосочка частота импульсации во вкусовых нервных волокнах в ответ на NaCl существенно снижается. Локальная фиксация потенциала одиночной вкусовой рецепторной клетки также показывает, что поток Na+ внутрь устраняется амилоридом. Поэтому и сделано заключение, что поток Na+ внутрь клетки вызывает деполяризацию вкусовой рецепторной клетки, приводящую к выделению медиатора на соседние нервные окончания.
Более глубокое понимание молекулярно-биологической природы амилорид-чувствительных Na+-каналов достигнуто с помощью клонирования гомологичных каналов из дистального отдела толстой кишки крысы. Этот ЭNaK состоит из трех субъединиц по 75 кДа. Использование антител к ЭNaK позволяет локализовать амилорид-чувствительные каналы вкусовых рецепторных клеток. Также показано, что ЭNaK присутствуют в эпителиальных клетках, окружающих вкусовые почки. Из чего предполагается, что в этих клетках существуют Na+-пути, модулирующие активность вкусовых рецепторных клеток. Тот факт, что ощущение солености детектируется не через G-белковую мембранную систему, а прямо через ионные каналы, объясняет, почему ответ на NaCl очень быстр. У крыс и хомячков латентный период ответа на NaCl составляет менее 50 мс (для сравнения – более 300 мс на сахарин). Ответ на соль также быстро адаптируется, часто менее, чем за 1 мин. Молекулярный механизм адаптации до сих пор неизвестен. Чувствительность к соленому находится под гормональным контролем. Внесение альдостерона в перфузионную жидкость кровеносного сосуда языка лягушки увеличивает ответ языкоглоточного нерва на аппликацию NaCl. Этот эффект сохраняется в течение 3-6 ч после перфузии. В других тканях показано, что альдостерон увеличивает вероятность открытого состояния предсуществующих Na+-каналов на срок около 10 ч, повышая, таким образом чувствительность. Действует ли такой же механизм и во вкусовой рецепторной клетке пока неизвестно. Другим гормоном, влияющим на чувствительность к соленому, является антидиуретический гормон (АДГ). Аппликация этого гормона также вызывает медленное усиление ответа языкоглоточного нерва на аппликацию соли на язык. АДГ выделяется гипофизом, когда осмолярность плазмы возрастает. Если это одновременно делает вкусовые рецепторные клетки более чувствительными к NaCl, животное, по-видимому, уменьшает потребление соленой пищи и, таким образом, компенсирует повышение осмолярности.
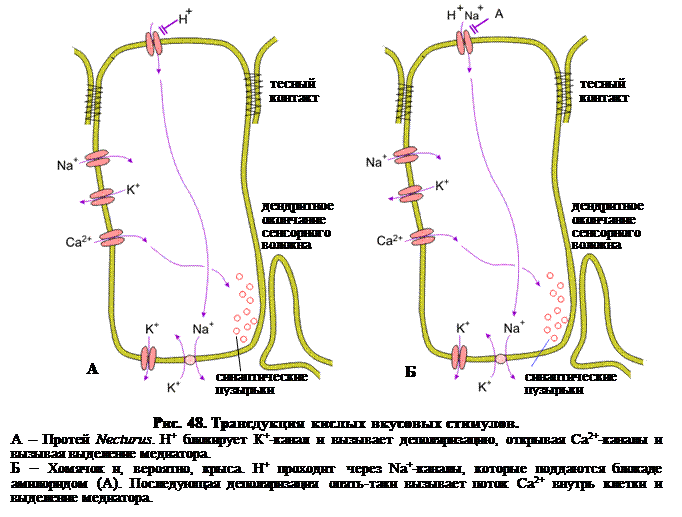
Кислое. Детектирование кислотности также имеет важное значение для поддержания гомеостаза внутренней среды. В этом случае воздействию подвергается рН внутренней среды. Детектирование кислого также важно для того, чтобы предупредить употребление в пищу неспелых (т.е. в питательном отношении неоптимальных) или скисших фруктов. Полагают, что кислый вкус обусловлен концентрацией свободных ионов Н+. Н+, как и Na+, вероятно влияет на каналы вкусовых рецепторных клеток непосредственно, причем у разных позвоночных обнаружено несколько различных механизмов такого действия. У земноводных, таких как протей Necturus, H+ блокирует каналы утечки К+ (рис. 48, А). Поскольку каналы утечки К+ играют существенную роль в поддержании потенциала покоя их блокада ведет к деполяризации. Это, в свою очередь, приводит к экзоцитозу медиатора из синаптических пузырьков. У хомячка и, вероятно, у крысы существует иной механизм. В этом случае (см. рис. 48, Б) Н+ проходит через амилорид-чувствительные Na+-кaналы. Этот поток катионов через вкусовые рецепторные клетки также вызывает деполяризацию и, следовательно, выделение медиатора.
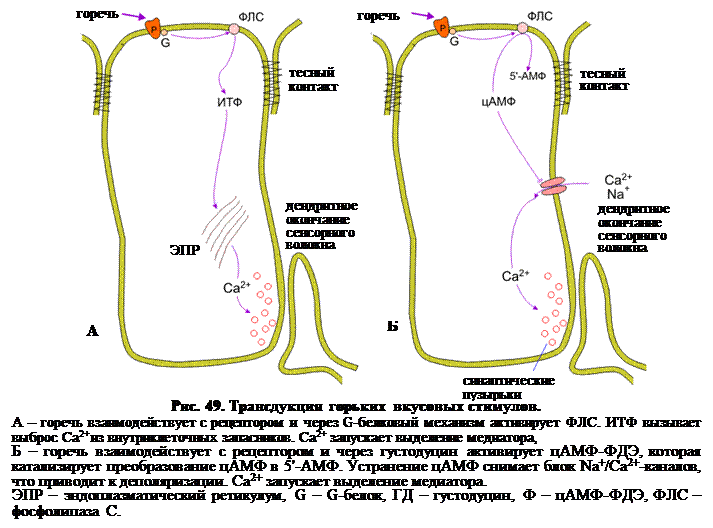
Горькое. Как и кислое, горечь вызывает реакцию отвращения. Имеющие горький вкус компоненты пищевых продуктов, например, растительные алкалоиды – хинин, стрихнин и т. д., цианид в горьком миндале, а также продукты распада, как например мочевина, отвергаются. Горький вкус имеют многие вещества, и возможно, что некоторые растения и беспозвоночные приобрели способность накапливать эти вещества с целью защиты. В некоторых случаях на основе этого наблюдается бэйтсовская мимикрия, когда организмы, не обладающие ядовитыми или неприятными веществами, выживают путем обмана, за счет подражания тем организмам, которые такие вещества содержат. Вероятно, и здесь существует более одного механизма трансдукции. Самым важными из них является блокада К+-каналов (хотя этот механизм мало избирателен) и более специфичные механизмы с участием системы G-белков. Во вкусовых рецепторных клетках крыс и мышей обнаружены два механизма (рис. 49): Ø взаимодействие горького вещества со специфическим рецептором приводит к активации мембранно-связанной фосфолипазы С и продукции ИТФ и ДАГ. ИТФ, как и обычно, вызывает выброс Са2+ из запасников в ЭР, а тот, в свою очередь, запускает экзоцитоз медиатора (рис. 49 А); Ø густодуцины (передатчики вкуса) активируют фосфодиэстеразу цАМФ (цАМФ-ФДЭ), которая катализирует преобразование цАМФ в 5-АМФ. При повышенной концентрации циклических нуклеотидов катионные каналы вкусовых рецепторных клеток не открываются, (что вызвало бы деполяризацию клетки) а, наоборот, закрываются, поддерживая высокий потенциал покоя. Гидролиз цАМФ под действием ФДЭ приводит к открыванию каналов для Na+ и Са2+ и деполяризации вкусовой клетки (рис. 49 Б).
Вода. Потребление адкватных количеств воды сущственно для поддержания соответствующей осмолярности жидкости в организме. Детекторы соленого, как описано выше, играют важную роль в поддержании этого баланса, одновременно демонстрируя и присутствие детекторов воды. Языкоглоточный нерв лягушки и смешанная ветвь лицевого нерва (chorda tympani) млекопитающих содержат волокна, отвечающие на аппликацию воды на язык, в то время как другие волокна реагируют на NaCl. Поскольку ацетат натрия не меняет реакцию на воду, тогда как NaCl и холин-хлорид влияют на нее, было сделано предположение, что этот механизм зависит от хлорных каналов в апикальной мембране. Снижение наружной концентрации хлорида активирует эти каналы; это приводит к повышению секреции С1-, что и ведет к деполяризации вкусовой рецепторной клетки. Как и в случае описанных выше механизмов восприятия вкусов, возможны и иные биохимические пути, исследование которых продолжаются. Субъективно «вкус» воды меняется в зависимости от того, какое вкусовое ощущение ему предшествовало. Если перед этим действовал хинин или лимонная кислота, вкус воды воспринимается, как сладкий, тогда как после NaCl или сахарозы – как горький. Из этого вытекает, что, возможно, между рецепторами воды и рецепторами упомянутых вкусовых ощущений существует взаимодействие.
Это ядро – сложное образование, получающее афференты из многих внутренних органов. Вкусовые волокна оканчиваются в узкой зоне, называемой вкусовым ядром. Отсюда нервные волокна восходят к мелким клеткам вентрального заднего ядра таламуса. Вкусовая информация затем передается в две области коры мозга – первая лежит в постцентральной извилине (поле Бродмана 3b), а вторая – глубоко в Сильвиевой борозде, на наружной поверхности инсулы (островковой доли). Обе они расположены вблизи от соматосенсорных зон языка.
До настоящего времени остается неясным вопрос о том, как мозг распознает вкусовые стимулы – по тому, какие группы волокон активируются, или по тому, какой характер ответа наблюдается во всей массе волокон. Существуют аргументы в пользу обоих механизмов, но на данный момент общее мнение склоняется в пользу последнего. Большинство фактов говорит в пользу широкой чувствительности, а не узкой специализации вкусовых клеток.
|

 Сладкое. Рецепторные клетки сладкого, как показывает рис. 46, имеют, по крайней мере два сигнальных пути. Сами по себе сахара, по-видимому, приводят в действие сигнальный путь с участием G-белков, активируя мембранный рецептор сахарозы. Есть данные, что это семидоменный рецептор, схожий с Са2+-чувствительным рецептором (СаЧР) и метаботропным глутаматными рецептором (mGluR). Этот рецепторный белок, известный как TR1, имеет очень длинную внеклеточную N-концевую цепь, которая, вероятно, и образует сайты вкусового рецептора. Когда TR1 активируется соответствующим вкусовым стимулом, G-белок сигнальная система вызывает выделение в цитозоль цАМФ, который через протеинкиназу А (ПКА) закрывает К+-канал. Циклический АМФ сам по себе также способен открывать Na+/Ca2+-вopoтный механизм. Результат действия этих двух механизмов – деполяризация. Это, в свою очередь, ведет к потоку Са2+ внутрь клетки, который запускает экзоцитоз медиатора из синаптических пузырьков. Несахарные подсластители, такие как сахарин, сладкие белки (тауматин и т. д.), некоторые аминокислоты и т. д. действуют иным путем. Существуют данные, что в этом случае стимулируется система фосфолипазы С (ФЛС), приводящей к выделению ИТФ (инозитол-три-фосфата) в цитозоль. Это ведет к выделению Са2+ из запасников в эндоплазматическом ретикулуме, что инициирует выделение медиатора из синаптических пузырьков.
Сладкое. Рецепторные клетки сладкого, как показывает рис. 46, имеют, по крайней мере два сигнальных пути. Сами по себе сахара, по-видимому, приводят в действие сигнальный путь с участием G-белков, активируя мембранный рецептор сахарозы. Есть данные, что это семидоменный рецептор, схожий с Са2+-чувствительным рецептором (СаЧР) и метаботропным глутаматными рецептором (mGluR). Этот рецепторный белок, известный как TR1, имеет очень длинную внеклеточную N-концевую цепь, которая, вероятно, и образует сайты вкусового рецептора. Когда TR1 активируется соответствующим вкусовым стимулом, G-белок сигнальная система вызывает выделение в цитозоль цАМФ, который через протеинкиназу А (ПКА) закрывает К+-канал. Циклический АМФ сам по себе также способен открывать Na+/Ca2+-вopoтный механизм. Результат действия этих двух механизмов – деполяризация. Это, в свою очередь, ведет к потоку Са2+ внутрь клетки, который запускает экзоцитоз медиатора из синаптических пузырьков. Несахарные подсластители, такие как сахарин, сладкие белки (тауматин и т. д.), некоторые аминокислоты и т. д. действуют иным путем. Существуют данные, что в этом случае стимулируется система фосфолипазы С (ФЛС), приводящей к выделению ИТФ (инозитол-три-фосфата) в цитозоль. Это ведет к выделению Са2+ из запасников в эндоплазматическом ретикулуме, что инициирует выделение медиатора из синаптических пузырьков.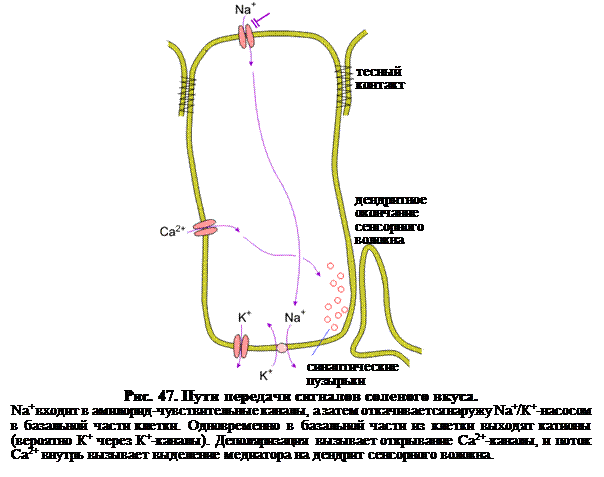
 Как отмечалось выше, вкусовые рецепторные клетки – это истинные рецепторные клетки. В ответ на деполяризацию они выбрасывают медиатор, действующий на дендритные окончания сенсорных волокон VII, IX и X черепно-мозговых нервов. Рис. 50 показывает, что волокна всех трех путей передачи вкусовых сигналов оканчиваются в ядре одиночного пучка продолговатого мозга.
Как отмечалось выше, вкусовые рецепторные клетки – это истинные рецепторные клетки. В ответ на деполяризацию они выбрасывают медиатор, действующий на дендритные окончания сенсорных волокон VII, IX и X черепно-мозговых нервов. Рис. 50 показывает, что волокна всех трех путей передачи вкусовых сигналов оканчиваются в ядре одиночного пучка продолговатого мозга.
 При отведении от смешанной ветви лицевого нерва хомячка, когда вкусовые почки подвергались воздействию различных концентраций одного и того же вкусового вещества, либо действию нескольких веществ одного вкуса, наблюдалась одна и та же (или очень похожая) картина ответа. Если концентрация вкусового вещества, например поваренной соли, меняется, суммарный ответ в 40 волокнах смешанной ветви лицевого нерва остается постоянным, хотя в каждом из одиночных волокон частота импульсов может повышаться или снижаться в зависимости от интенсивности стимула. Иными словами, тип активности может изменяться, но общий ее «профиль» остается неизменным. Аналогичная картина наблюдается для ряда групп веществ, вызывающих одинаковые (или схожие) вкусовые ощущения – для вещества сладкого вкуса (как сахарин, сахароза, фруктоза); для солей натрия (NaCl и NaNO3), и кислот (соляной и лимонной). Это электрофизиологическое наблюдение подкрепляется поведенческими экспериментами, показывающими, что хомячок не различает разные сладкие вещества и разные соли.
При отведении от смешанной ветви лицевого нерва хомячка, когда вкусовые почки подвергались воздействию различных концентраций одного и того же вкусового вещества, либо действию нескольких веществ одного вкуса, наблюдалась одна и та же (или очень похожая) картина ответа. Если концентрация вкусового вещества, например поваренной соли, меняется, суммарный ответ в 40 волокнах смешанной ветви лицевого нерва остается постоянным, хотя в каждом из одиночных волокон частота импульсов может повышаться или снижаться в зависимости от интенсивности стимула. Иными словами, тип активности может изменяться, но общий ее «профиль» остается неизменным. Аналогичная картина наблюдается для ряда групп веществ, вызывающих одинаковые (или схожие) вкусовые ощущения – для вещества сладкого вкуса (как сахарин, сахароза, фруктоза); для солей натрия (NaCl и NaNO3), и кислот (соляной и лимонной). Это электрофизиологическое наблюдение подкрепляется поведенческими экспериментами, показывающими, что хомячок не различает разные сладкие вещества и разные соли.


