Предмет, задачи и цели молекулярной биологии
The blue-green algae use chlorophyll a in photosynthesis, in combination with essentially the same carotenoid pigments and electron carriers found in eukaryotes. As in eukaryotes, the pigments are organized into two photosystems interconnected by the Z-pathway. Phycobilins (see Fig. 8-10), acting as accessory pigments, are also present as light-absorbing molecules. These unusual pigments are found in only one other group of photosynthetic organisms, the red algae. The
Figure 8-30 Flattened, elongated thylakoids in a purple photosynthetic bacterium, x 36, 000. Courtesy of W. C. Trentini, Mt. Allison University, Sackville, New Brunswick, Canada.
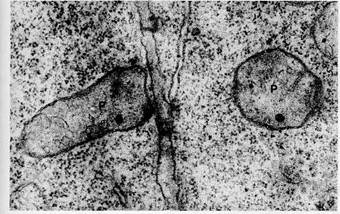
Figure 8-29 Spherical thylakoids in the cytoplasm of the purple photosynthetic bacterium Rhodopseudomonas. x 43, 000. From Biochemistry of Chloroplasts, Vol. 1, ed.T. W. Goodwin, 1966. Courtesy of W. Menke and Academic Press, Inc. presence of phycobilins in only the blue-green and red algae has led to much speculation about possible evolutionary relationships between the two groups. The photosynthetic apparatus in blue-green algae is bound to saclike membranes suspended directly in the cytoplasm, as in the bacteria (see Figs. 8-31 and 1-2). Attached to the outer surfaces of the sacs are spherical particles that appear slightly denser than the surrounding cytoplasm (arrows in Fig. 8-31). Dense particles of the same dimensions also appear in association with the thylakoids in red algae chloroplasts. Re-ce'ntly, E. Gantt and her coworkers (Gantt and Lipschultz, 1972) showed that these particles, when isolated, contain phycobilin pigments combined with proteins to form phyco-biliproteins (see pp. 173). The particles, called phycobilisomes, are believed to act as light-harvesting antennae associated. with photosystem II, acting in a manner analogous to the LHCP complex of chloroplasts in higher plants. For Further Information DNA, ribosomes, and protein synthesis in chloroplasts, Supplements 12-2 and 13-1 Evolutionary origins of chloroplasts, Chapter 17 Mitchell's chemiosmotic hypothesis, ^ Chapter 7 Peroxisomes and photorespiration, Supplement 7-3 Figure 8-31 Thylakoids in the cytoplasm of the blue-green alga Synechococcus. Numerous phycobilisomes (arrows) are attached to the thylakoid membranes, x 30, 000. Courtesy of M. R. Edwards, New York State Department of Health, from J. Cell Biol. 50: 896 (1971), by copyright permission of the Rockefeller University Press. Suggestions for Further Reading Anderson, J. M. 1977. The molecular organization of chloro-plast thylakoids. In International cell biology 1976-1977, ed. B. R. Brinkley and K. R. Porter, pp. 183-192. New York: Rockefeller University Press. Avron, M. 1977. Energy transductions in chloroplasts. Annu. Rev. Biochem. 46: 143-155. Blankenship, R. E., and Parson, W. W. 1978. The photochemical electron transfer reactions of photosynthetic bacteria and plants. Annu. Rev. Biochem. 47: 635-653. Carr, N. G., and Whitton, B. A. 1973. The biology of blue-green algae. Berkeley: University of California Press. Clayton, R. K. 1970. Light and living matter. 2 vols. New York: McGraw-Hill. Govindjee. 1975. Bioenergetics of photosynthesis. New York: Academic Press. Govindjee, and Govindjee, R. 1974. The primary events of photosynthesis. Sri. Amer. 231: 68-82. Halliwell, B. 1978. The chloroplast at work. A review of modern developments in our understanding of chloroplast metabolism. Prog. Biophys. Molec. Biol. 33: 1-54. Hatch, M. D. 1976. Photosynthesis: the path of carbon. In. Plant biochemistry, vol. 3, ed. J. Bonner and J. E. Varner, pp. 797-884. New York: Academic Press. Heber, V. 1974. Metabolite exchange between chloroplasts and cytoplasm. Annu. Rev. Plant Physiol. 25: 393-421. Horton, P. 1976. Organization and function of chloroplast photosystems. Internat. ]. Biochem. 7: 597-603. Kirk, J. T. O., and Tilney-Bassett, R. A. E. 1978. The plastids, 2nd ed. New York: Elsevier/North-Holland. Kok, B. 1976. Photosynthesis: The path of energy. In Plant biochemistry, vol. 3, ed. J. Bonner and J. E. Varner, pp. 845-885. New York: Academic Press. Park, R. B. 1976. The chlofoplast. In Plant biochemistry, vol. 3, ed. J. Bonner and J. E. Varner, pp. 115-145. New York: Academic Press. Parson, W. W. 1974. Bacterial photosynthesis. Annu. Rev. Microbiol. 28: 41-59. Trebst, A. 1974. Energy conservation in photosynthetic electron transport. Annu. Rev. Plant Phys. 25: 423-458. Whatley, F. R. 1975. Chloroplasts. Ciba Foundation Sym. 31: 41-104. Supplement 8-1: Plastids and Plastid Development Mature cells may contain a variety of organelles that are closely related to chloroplasts. These organelles, collectively called plastids, include as major types amyloplasts, etioplasts, and chromoplasts, as well as chloroplasts. Internally the stroma is specialized and distinctly different in each plastid type. In amyloplasts, which are colorless, the stroma is completely filled with one or more starch grains. Otherwise, the stroma is completely structureless. These plastids function as starch storage vesicles in roots and other tissues in plants. The stroma of chromoplasts, responsible for the red, yellow, or orange color of fruits, flower petals, and roots such as the carrot, contains deposits of carotenoid pigments, usually in the form of small granules. As much as 50% or more of the total weight of chromoplasts may consist of carotenoids. Etioplasts, found only in leaves growing in continuous darkness, contain a regular lattice of internal membranes and fibrous deposits of proteins. Etioplasts are capable of rapid differentiation into fully functional chloroplasts if dark-grown leaves are exposed to light. All of these plastid types develop from proplastids (Fig. 8-32), small bodies in which a relatively undifferentiated stroma is surrounded by inner and outer boundary membranes. A few tubular imaginations of the inner boundary membrane may extend into the stroma of proplastids. Fibrous DNA deposits, ribosomes, starch grains, and osmiophilic granules may also be present. Occasionally a limited number of single thylakoids may be scattered in the stroma. About 10-20 proplastids occur in meristematic cells of a growing stem; somewhat larger numbers, up to 40, are found in root meristems. In stems and leaves grown in the light, these bodies quickly develop into chloroplasts. Transformation of proplastids into chloroplasts occurs first by tubelike growths of the inner boundary membrane that gradually extend into the stroma and flatten. At points, the flat, thylakoidlike vesicles become doubly and triply folded into multiple layers (Fig. 8-33). This process repeats until typical grana are formed. Synthesis of chlorophyll and accessory pigments takes place during growth of the inner membranes. If plants grow in the dark, proplastids in leaves and stems develop into etioplasts instead of chloroplasts. These plastids are larger and more complex internally than proplastids and contain a characteristic lattice of internal membranes (Fig. 8-34). The lattices, called prolamellar bodies, are formed from a regular network of connected, membranous tubules. DNA deposits and ribosomes may be scattered in the stroma around the prolamellar bodies. Under the light microscope, etioplasts appear yellow, primarily because of carotenoid pigments, which may be present in quantities amounting to as much as one-third of the carotenoid content of mature chloroplasts. Also present is pro-tochlorophyll, a molecule identical to chlorophyll a except for the absence of two hydrogens in the porphyrin ring (see Fig. 8-8). These pigments are concentrated in the membranes of the prolamellar body, which also contain most of the protein components of photosystems I and II and the electron carriers linking them. Although most of the components necessary for photosynthesis are present, the photosystems and the electron transport system in etioplasts are inactive.
Figure 8-32 Proplastids (P) in the cytoplasm of adjacent tobacco cells, x 28, 000. Courtesy of*D. von Wettstein. Figure 8-33 An early stage in the development of a chloroplast in the leaf meristem of Oenothera. Initial folding of the membranes in the region of future grana stacks (arrows) has occurred, x 34, 000. Courtesy of W. Menke.
Figure 8-34 An etioplast from an Avena leaf. A highly ordered prolamellar body is present in the matrix; also visible are numerous ribosomes (arrows), x 44, 000. Courtesy of B. E. S. Gunning, from Protoplasma 60: 111 (1965).
Figure 8-35 Development of thylakoids from the prolamellar body in a greening chloroplast from an oat leaf. Courtesy of J. T. 0. Kirk. When etiolated plants are placed in light, the leaves rapidly turn green. The wavelengths of light effective in inducing the transformation of etioplasts are those absorbed by pro-tochlorophyll, which, under the influence of light, is rapidly converted to chlorophyll a. This conversion may be completed in as little as 1 min after exposure to light. Appearance of chlorophyll b is much slower; this pigment cannot be detected until several hours after transfer of plants to light. As protochlorophyll is converted to chlorophyll a, the tubules of the prolamellar body are gradually transformed to thylakoids. The details of this process have been studied by D. von Wettstein (1958) and B.E.S. Gunning (1965). If the light intensity is high, the tubules flatten within minutes into thy- lakoidlike sacs that extend into the stroma of the developing chloroplast (Fig. 8-35). The developing thylakoids gradually layer into stacks at scattered points by a process similar to grana formation in proplastids. This process repeats until typical grana form. Completion of membrane folding transforms the etioplasts into typical chloroplasts. These changes are correlated with the appearance of full biochemical activity. Within 30 min after etioplasts begin to green, photosystem I activity is detectable. Somewhat later, within a matter of hours, photosystem II becomes active. Although formation of grana stacks is not required for photosystem II activity, appearance of the biochemical activity of this system is usually correlated with grana development. Предмет, задачи и цели молекулярной биологии. Возникла во второй половине 20 в. Название этой науки связано с именем У. Эстбери, который в 39 г. назвал себя молекулярным биологом, через 2-а года он же получил 1-ую рентгенограмму ДНК и тем самым положил начало самой главной молекулы, впервые выявленные Мишиным. Первое официальное упоминание о молекулярной биологии принадлежит У. Уиверу, руководителю Рокфеллеровского фонда, в 1938 г. написал: «В тех пограничных областях, где химия и физика перекликается с биологией, постепенно возникает новый раздел науки – молекулярная биология, начинающая приоткрывать завесы над многими тайнами, окутывающими основные элементы живой природы». Т. о. было постулировано возникновение нового направления современной биологии, которое интегрировало усилиями в области объектов живой природы. Центром молекулярно-биологических исследований стали работы в области изучения материальных основ наследственности природы генов и механизмов передач, наследование признаков из поколения в поколение. Важнейшие направления молекулярной биологии: -Молекулярная генетика – исследование структурно-функциональной организации генетического аппарата клетки и механизма реализации наследственной информации. -Молекулярная вирусология – исследование молекулярных механизмов взаимодействия вирусов с клетками. -Молекулярная иммунология – изучение закономерностей иммунных реакций организма. -Молекулярная биология развития – исследование появления разнокачественности клеток в ходе индивидуального развития организмов и специализации клеток.
|