Гистология
Кора и центральные ядра мозжечка образуются серым веществом, остальная масса — белым. На поперечных срезах коры мозжечка (рис. 6) различают поверхностный, или молекулярный, слой (stratum moleculare) и глубокий, или зернистый, слой (stratum granulosum), между которыми расположен узкий ганглиозный слой (stratum gangliosum), образованный одним рядом клеток Пуркинье очень больших размеров. От основания клетки Пуркинье отходит осевоцилиндрический отросток, проникающий через слой зерен в белое вещество. При этом почти от начальной части отростка отходят коллатерали, заканчивающиеся частично в слое зерен, частично около соседних клеток Пуркинье. От верхнего конца клетки Пуркинье в молекулярный слой отходят разветвляющиеся дендриты, расположенные всегда в одной плоскости, перпендикулярно к длине извилины. Помимо этих разветвлений, в молекулярном слое расположены маленькие мультиполярные клетки с коротким ветвящимся аксоном и большие, «корзинчатые», клетки с длинным аксоном, разветвления которого заканчиваются в виде сплетений («корзинок») около многих клеток Пуркинье. В зернистом слое расположены очень маленькие клетки-зерна, дендриты которых окружают соседние клетки и входят в соприкосновение с различными нервными волокнами, а осевоцилиндрические отростки направляются в восходящем направлении в молекулярный слой и разветвления их заканчиваются пуговками на разветвлениях дендритов многих клеток Пуркинье и «корзинчатых» клеток. Среди клеток-зерен в зернистом слое встречаются в небольшом количестве звездчатые клетки (клетки Гольджи), дендриты которых разветвляются в зернистом и молекулярном слоях и в слое клеток Пуркинье, короткие аксоны разветвляются в слое зерен, а длинные аксоны уходят в белое вещество. Афферентная система мозжечка образована мшистыми и ползучими волокнами, направляющимися из ножек мозжечка через белое вещество в кору мозжечка. Клетки Пуркинье получают импульсы непосредственно через мшистые и ползучие волокна или посредством клеток-зерен. Клетки-зерна получают импульсы от мшистых волокон и передают их через свои аксоны в молекулярный слой на дендриты «корзинчатых» и звездчатых клеток, а звездчатые клетки через свои аксоны посылают импульсы к клеткам Пуркинье. Кроме того, в системе многочисленных разветвлений мшистых волокон, клеток-зерен и звездчатых клеток имеются и замкнутые круги действия.
В вегетативной нервной системе различают центральные и периферические отделы. Центральные отделы симпатической нервной системы представлены ядрами боковых рогов тораколюмбального отдела спинного мозга. В парасимпатической нервной системе центральные отделы включают ядра среднего и продолговатого мозга, а также ядра боковых рогов сакрального отдела спинного мозга. Парасимпатические волокна краниобульбарного отдела выходят в составе III-й, VII-й, IX-й и Х-й пар черепных нервов. Вегетативные рефлекторные дуги начинаются чувствительным нейроном, тело которого лежит в спинномозговом узле (ганглии), как и в соматической рефлекторной дуг. Ассоциативные нейроны находятся в боковых рогах спинного мозга. Здесь нервные импульсы переключаются на промежуточные преганглионарные нейроны, отростки которых покидают центральные ядра и достигают вегетативных ганглиев, где передают импульсы на двигательный нейрон. В связи с этим различают нервные волокна преганглионарные и постганглионарные. Первые из них покидают центральную нервную систему в составе вентральных корешков спинномозговых нервов и черепных нервов. Как в симпатической, так и в парасимпатической системах преганглионарные нервные волокна принадлежат холинергическим нейронам. Аксоны нейронов, расположенных в вегетативных ганглиях, называются постганглионарными. Они не образуют прямых контактов с эффекторными клетками. Их терминальные отделы по своему ходу формируют расширения — варикозности, в составе которых находяся пузырьки медиатора. В области варикозности нет глиальной оболочки и нейромедиатор, выделяясь в окружающую среду, влияет на эффекторные клетки (например, на клетки желез, гладкие миоциты и др.). В периферических ганглиях симпатической нервной системы, как правило, находятся адренергические эфферентные нейроны (за исключением нейронов, имеющих синаптические связи с потовыми железами, где симпатические нейроны являются холинергическими). В парасимпатических ганглиях эфферентные нейроны всегда холинергические.
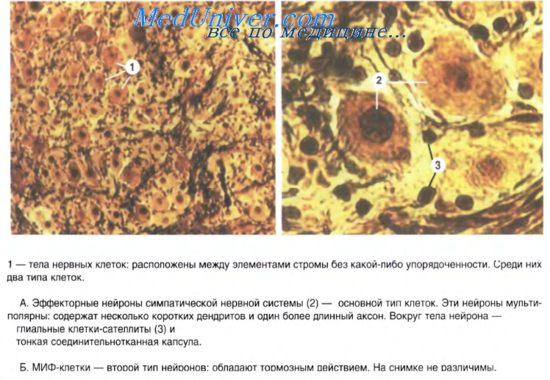
Ганглии представляют собой скопления мультиполярных нейронов (от нескольких клеток до десятков тысяч). Экстраорганные (симпатические) ганглии имеют хорошо выраженную соединительнотканную капсулу, как продолжение периневрия. Парасимпатические ганглии находятся, как правило, в интрамуральных нервных сплетениях. Ганглии интрамуральных сплетений, как и другие вегетативные узлы, содержат вегетативные нейроны местных рефлекторных дуг. Мультиполярные нейроны диаметром 20-35 мкм расположены диффузно, каждый нейрон окружен глиоцитами ганглия. Кроме того, описаны нейроэндокринные, хеморецепторные, биполярные, а у некоторых позвоночных и униполярные нейроны. В симпатических ганглиях имеются мелкие интенсивно флюоресцирующие клетки (МИФ-клетки) с короткими отростками и большим количеством гранулярных пузырьков в цитоплазме. Они выделяют катехоламины и оказывают тормозящее влияние на передачу импульсов с преганглионарных нервных волокон на эфферентный симпатический нейрон. Эти клетки называют интернейронами. Среди крупных мультиполярных нейронов вегетативных ганглиев различают: двигательные (клетки Догеля 1-го типа), чувствительные (клетки Догеля П-го типа) и ассоциативные (клетки Догеля Ш-го типа). Двигательные нейроны имеют короткие дендриты с пластинчатыми расширениями ("рецептивные площадки"). Аксон этих клеток очень длинный, уходит за пределы ганглия в составе постганглионарных тонких безмиелиновых нервных волокон и оканчивается на гладких миоцитах внутренних органов. Клетки 1-го типа называют длинноаксонными нейронами. Нейроны П-го типа — равноотростчатые нервные клетки. От их тела отходят 2-4 отростка, среди которых различить аксон трудно. Не разветвляясь, отростки уходят далеко от тела нейрона. Их дендриты имеют чувствительные нервные окончания, а аксон оканчивается на телах двигательных нейронов в соседних ганглиях. Клетки П-го типа являются чувствительными нейронами местных вегетативных рефлекторных дуг. Клетки Догеля Ш-го типа по форме тела похожи на вегетативные нейроны П-го типа, но их дендриты не выходят за пределы ганглия, а нейрит направляется в другие ганглии. Многие исследователи считают эти клетки разновидностями чувствительных нейронов. Таким образом, в периферических вегетативных ганглиях имеются местные рефлекторные дуги, состоящие из чувствительных, двигательных и, возможно, ассоциативных вегетативных нейронов. Интрамуральные вегетативные ганглии в стенке пищеварительного тракта отличаются тем, что в их составе, кроме двигательных холинергических нейронов, имеются тормозные нейроны. Они представлены адренергическими и пуринергическими нервными клетками. В последних медиатором является пуриновый нуклеотид. В интрамуральных вегетативных ганглиях встречаются также пептидергические нейроны, выделяющие вазоинтестинальный пептид, соматостатин и ряд других пептидов, с помощью которых осуществляются нейроэндокринная регуляция и модуляция деятельности тканей и органов пищеварительной системы. Основополагающие открытия в области физиологии, гистологии, фармакологии были сделаны выдающимися талантливыми учеными России начиная с середины XIX в. Им принадлежал приоритет в работах по изучению роли ЦНС в иннервации сердца, морфологического строения периферического отдела вегетативной нервной системы сердца и других органов, а также в исследованиях влияния фармакологических веществ - атропина, мускарина, никотина, хлороформа на функцию сердца. Профессор Киевского университета Александр Петрович Вальтер (1817-1889) в 1843 г, впервые доказал, что раздражение симпатического нерва вызывает сужение кровеносных сосудов (в опытах на кролике) [1]. Петербургский ученый Илья Фаддеевич Цион (1842-1912) в 1866 г. в лаборатории К.Людвига открыл ускоряющее влияние симпатического нерва на сердце и обнаружил центростремительный (афферентный) нерв, начинающийся от дуги аорты, который рефлекторно понижал артериальное давление, если аорта растягивалась вследствие повышения артериального давления в ней [7]. Основоположник казанской школы физиологов Филипп Васильевич Овсянников (1827-1906), уже работая в Петербургском университете, в 1871 г. открыл сосудодвигателъный центр в продолговатом мозге и установил, что симпатический и парасимпатический нервы оказывают противоположное влияние на сердце и другие органы [5]. Основатель казанской школы фармакологов Иван Михайлович Догель (1830-1916} в 1866 г. впервые вызвал рефлекторную остановку сердца при раздражении окончаний чувствительных нервов верхних дыхательных путей хлороформом [10]. Владимир Васильевич Николаев, талантливый представитель казанской школы физиологов и фармакологов, сделал в 1893-1894 гг. открытие о двухнейронном строении сердечных ветвей блуждающего (парасимпатического) нерва [3, 11]. В.В. Николаев родился 17 февраля 1871 г. I в г. Задонске Воронежской губернии в семье служащего. После окончания в 1890 г. Воронежской гимназии поступил на медицинский факультет Казанского университета, который окончил в 1895 г. и получил диплом лекаря с отличием. Студентом он начал работать в научном студенческом кружке при физиологической лаборатории, которой руководил выдающийся русский физиолог профессор Николай Александрович Миславский (1854- 1929). Под его руководством проводились работы по изучению нервных окончаний в сердце, ответственных за реализацию тормозных и возбуждающих импульсов, исходящих из ЦНС. К тому времени было известно, что в иннервации сердца участвуют симпатический и блуждающий нервы. Симпатический нерв на своем пути из ЦНС к сердцу прерывался на нервных узлах (на современном языке эти узлы называются симпатическими ганглиями). Сердечные же ветви блуждающего нерва при выходе из продолговатого мозга никаких нервных узлов по ходу к сердцу не имели. Перед исследователями встал вопрос: есть ли подобные узлы или нервные клетки у блуждающего нерва и где они располагаются в сердце? Русский физиолог и гистолог профессор Юрьевского университета Фридрих Генрих Биддер (1810-1894) еще в 50-х гг. XIX в. наблюдал в сердце лягушки целую сеть сплетающихся нервных волокон, часть из которых оканчивалась не на мышечных элементах, а на неких нервных ("ганглиозных") клетках, залегающих в толще миокарда. Установить принадлежность этих клеток к симпатическому или блуждающему нерву в то время было невозможно, и Ф.Г. Биддер выдвинул лишь гипотезу, что эти нервные клетки могли принадлежать к блуждающему нерву [8,9]. Н.А. Миславский предложил выяснить этот вопрос В.В. Николаеву. В 1893-1894 гг. 22-летний студент медицинского факультета В.В. Николаев поставил на сердце лягушки длительный и многоплановый эксперимент, требовавший ювелирной техники исполнения. Он применил метод окраски нервных образований в сердце лягушки метиленовой синью и осмиевой кислотой. Наблюдая под микроскопом нервные клетки и их окончания, молодой ученый подтвердил результаты, полученные Ф.Г. Биддером, и исследовал взаимоотношения нервных клеток округлой формы и их двух отростков, один из которых был длинный и прямой, а другой - короткий в виде спирали. Эти отростки заканчивались нервными окончаниями. Чтобы доказать принадлежность этих клеток к блуждающему нерву, В.В. Николаев перерезал сердечные ветви вагуса и оставил жить лягушек на срок от 14 до 73 дней. На протяжении этого времени он наблюдал перерождение окончаний нерва на клетках округлой формы. Спиральные отростки, контактирующие с округлой нервной клеткой, перерождались, и на их месте образовывались капельки жира, окрашенные в черный цвет осмиевой кислотой. К концу 73-го дня после ваготомии спиральные окончания нерва разрушились, переродились, перестали существовать, а нервные клетки округлой формы сохранились. Это морфологическое наблюдение позволило В.В. Николаеву сделать вывод, что нервные округлые ("ганглиозные") клетки, обнаруженные Ф.Г. Биддером в сердце лягушки, принадлежали к блуждающему нерву. Так студент медицинского факультета Казанского университета В.В. Николаев сделал открытие о том, что блуждающий (парасимпатический) нерв сердца имеет двухнейронное строение. Тело первого парасимпатического нейрона (блуждающего нерва) находится в продолговатом мозге, а тело второго парасимпатического нейрона - в толще миокарда, на поверхности кардиомиоцита. Результаты этой работы В.В. Николаев доложил на заседании Казанского общества невропатологов. Статья "Об иннервации сердца лягушки" в 1893 г. была напечатана в немецком журнале [11], а в 1894 г. - в "Неврологическом вестнике" [3]. Весной 1895 г. после окончания университета В.В. Николаев был приглашен ассистентом на кафедру фармакологии Казанского университета, которую возглавлял тогда профессор И.М. Догель. Под его руководством В.В. Николаев выполнил оригинальную диссертацию на тему "Фотографирование глазного дна животных", которую успешно защитил в 1902 г. В том же году он был избран на должность приват-доцента по кафедре фармакологии и приступил к чтению лекций студентам. К тому времени В.В. Николаев проявил выдающиеся способности ученого и педагога. Поэтому ученый совет Казанского университета рекомендовал направить его за границу для дальнейшего усовершенствования в области научных исследований и изучения педагогических систем в различных университетах. В.В. Николаев с 1903 по 1906 г. слушает лекции в университетах Германии и Франции: по фармакологии у проф. О.Шмидеберга, по биологической химии у проф. Гофмейстера, по фармакогнозии у проф. Шера, по органической химии у проф. Фауста, по качественному анализу лекарственных средств у доктора Габейна. Изучение этих взаимосвязанных сложных предметов позволило ему приобрести широкие знания и освоить комплексную методологию научных исследований и построения преподавания, стать энциклопедически образованным ученым и поставило его в ряд выдающихся специалистов по фармакологии и фармации в России и Европе. В.В. Николаев во время своего пребывания в Страсбурге у проф. О.Шмидеберга в 1903-1906 гг. возвратился к фармакологическим опытам на сердце лягушки. Он изучал влияние мускарина и атропина на сердце лягушки с перерожденными окончаниями блуждающего нерва. Эти опыты были одними из первых в мировой науке по изучению действия веществ на органы животных с экспериментальной патологией. В этих опытах казанский ученый показал, что мускарин и атропин в условиях перерождения окончаний блуждающего нерва полностью сохранили свое действие на сердце. Ранее считалось, что атропин и мускарин изменяли работу сердца благодаря влиянию на окончания блуждающего нерва. Но в опытах окончания блуждающего нерва подверглись жировому перерождению и не могли реагировать на мускарин и атропин. В.В. Николаев делает вывод, что в сердце лягушки есть "самостоятельный аппарат", присутствия которого достаточно, чтобы получить замедление или остановку сердца от мускарина и увеличение числа сердечных сокращений от атропина. Истоки представления об этом "самостоятельном аппарате" стали формироваться лишь в 20-х гг. XX в. в связи с развитием теории о рецепторах. В первой половине XX в. возникло учение об М - холинорецепторах миокарда, возбуждение которых мускарином и ацетилхолином вызывает торможение сердца, а блокада их атропином - учащение работы сердца. Результаты этого исследования были опубликованы в 1910 г. [4] и в дальнейшем положили начало новому направлению по исследованию парасимпатической иннервации сердца у теплокровных. Так, в начале 20-х гг. XX в. Иван Петрович Разенков (1888-1954) провел эксперименты на кошках и собаках и подтвердил двухнейронное строение парасимпатического нерва [6]. Детальное гистологическое изучение парасимпатических ганглиев сердца было проведено Борисом Иннокентьевичем Лаврентьевым (1892-1944) [2]. В.В. Николаев по возвращении из-за границы, с 1909 г. читал доцентский курс, а с 1915 по 1921 г. заведовал кафедрой фармации и фармакогнозии Казанского университета. 19 сентября 1921 г. его избирают заведующим кафедрой фармакологии в Московском университете и одновременно директором Фармакологического института при кафедре. С 1930 г. (после образования I Московского медицинского института им. И.М. Сеченова) он являлся заведующим кафедрой фармакологии с рецептурой. В 1934 г. Высшая квалификационная комиссия Наркомздрава присудила ему ученую степень доктора медицинских наук, а в 1940 г. за выдающиеся заслуги в области медицины ему было присвоено звание "Заслуженный деятель науки РСФСР". Вся дальнейшая жизнь В.В. Николаева была связана с I ММИ им. И.М. Сеченова, где он проработал до 1950 г. Обладая энциклопедическими знаниями по фармакологии, фармакогнозии, химии, фармации, В.В. Николаев создает научный фундамент учения о лекарственных травах. Он берет на себя руководство фармакологическим отделом Всесоюзного института лекарственных и ароматических растений и организует фармакологические исследования многих растений. Например, в этот период им были открыты седативные свойства синюхи, кардиотоническое действие желтушника. Он предложил для клинического применения препарат из желтушника "Эризид". До сих пор отечественная промышленность выпускает препарат "Кардиовален", в состав которого входят гликозиды желтушника, впервые изученные В.В. Николаевым. Одновременно он изучает фармакологические свойства многих препаратов - хлороформа, атропина, мускарина, никотина бромида и других веществ. Им опубликовано свыше 130 статей в научных журналах и сборниках. В.В. Николаев написал 350 статей в различные справочные издания по фармакологии, фармации и медицине. По своему содержанию это были научные официозные статьи, среди них "Список ядовитых и сильнодействующих веществ" для Государственной Фармакопеи, свыше 200 статей для Малого медицинского энциклопедического словаря (1924), 59 статей для Большой медицинской энциклопедии (1928-1935), 17 статей для Большой советской энциклопедии (1926-1947). В.В. Николаев руководил научными работами многих аспирантов, 20 из которых защитили диссертации. 14 его учеников стали профессорами и заведующими кафедрами в Казани, Перми, Москве, Архангельске, Ереване, Львове. Это профессора В.М. Соколов, Н.Н. Кампанцев, Л.И. Медведев, А.Б. Вольтер, К.В. Бенинг, М.А. Левченко, Л.Д. Турова, Н.С. Правдин, В.М. Наумов, А.Н. Кудрин. С 1895 по 1950 г., т.е. 55 лет В.В. Николаев занимался преподаванием фармакологии, фармакогнозии, фармации на медицинских и фармацевтических факультетах. Он ввел в систему обучения целый ряд усовершенствований. На лекциях и практических занятиях он показывал опыты на экспериментальных животных, демонстрировавшие действие лекарственных веществ в зависимости от лекарственной формы, способа введения, состояния организма, фармацевтических факторов. Это направление явилось истоком для формирования современной биофармации. В.В. Николаев был одним из организаторов высшего медицинского и фармацевтического образования в России и СССР. Он участвовал в организации Казанского женского медицинского института, кафедр фармакологии в Смоленском (1923), III Московском медицинском (1934), Московском фармацевтическом (1935) институтах, в последнем был заведующим до 1937 г. Во всех институтах он читал лекции по фармакологии, организовывал педагогический процесс и готовил педагогические кадры. Он был председателем 12 государственных экзаменационных комиссий по выпуску врачей и провизоров. За свою 55-летнюю деятельность он подготовил огромную армию провизоров (фармацевтов) и врачей. Большая работоспособность, постоянная инициатива, стремление отдать все силы и знания народу были основными чертами личности В.В. Николаева. Это позволяло ему вести большую общественную деятельность. В 1895 г. он был ученым секретарем Казанского общества невропатологов, с 1896 г. - членом Казанского общества врачей, в 1905 г. участвовал в организации Казанского народного университета, в 1919 г. организовал и возглавил Казанское научное общество фармацевтов, в 1929 г. организовал Московское научное общество фармацевтов и был его председателем до 1925 г. В.В. Николаев был соавтором и редактором Государственной Фармакопеи 7-го издания, ответственным редактором журнала "Фармация и фармакология", членом редакционного совета "Физиологического журнала", членом физиологического общества им. И.М. Сеченова. Он выступал с научными проблемными докладами на съездах физиологов, фармакологов, биохимиков. В 1946 г. за 50 летнюю научную и педагогическую работу Президиум Верховного Совета СССР наградил В.В. Николаева орденом Трудового Красного Знамени. В.В. Николаев выполнил громадную работу по созданию советской медицинской и фармацевтической науки, высшего образования, медицинской промышленности, научных обществ. Когда знакомишься с разносторонними и глубокими по своему содержанию трудами В.В. Николаева, невольно поражаешься глубиной научной мысли, логической аргументацией выводов, способностью определять пути дальнейших научных исследований и внедрения их результатов в практическую медицину и фармацию. Его открытия в физиологии, фармакологии и фармации не были плодом счастливых случайностей, а представляли собой результат теоретических, научных осмыслений, оригинально спланированного и талантливо проведенного эксперимента. Залогом столь значительных успехов, несомненно, была природная одаренность В.В. Николаева, его громадный труд и удивительно благодатная атмосфера в Казанском университете, где в ту пору работали выдающиеся ученые, обеспечившие российский ренессанс в биологической и медицинской науках. В московский период работы у В.В. Николаева раскрылись новые таланты по организационному становлению фармакологии, фармации и медицины. Его труды были основой для формирования фармацевтической промышленности, оценки качества лекарств. Труды В.В. Николаева в области лекарственных растений явились фундаментом для научной фитотерапии. Залогом успеха В.В. Николаева, помимо одаренности и энциклопедической образованности, были любовь к Родине, ответственность, организованность, скромность и трудолюбие. В.В. Николаев прожил долгую жизнь. Он скончался в Москве в 1950 г. в возрасте 79 лет.
22. Роговица, склера, хрусталик. Стекловидное тело глазного яблока. Развитие глаза.
|