Еще в далекие 70-е годы мы получили косвенные доказательства того, что синаптонемный комплекс формируется путем самосборки его элементов, которая может происходить и в отсутствие хромосом. Эксперимент поставила сама природа, а нам удалось его наблюдать. Оказалось, что у свиной аскариды в цитоплазме клеток, готовящихся к мейозу I, появляются пакеты или «штабеля» абсолютно правильно уложенных морфологических элементов синаптонемного комплекса (хотя в цитоплазме нет хромосом: они — в ядре). Поскольку на стадии подготовки клеток к мейозу в клеточных ядрах еще нет синаптонемного комплекса, появилось предположение о несовершенстве контроля очередности событий мейоза у этого примитивного организма. Избыток новосинтезированных белков в цитоплазме приводит к их полимеризации и возникновению структуры, не отличающейся от синаптонемного комплекса [3]. Эта гипотеза получила подтверждение только в 2005 г. благодаря работе интернациональной группы исследователей, работающих в Германии и Швеции. Они показали, что если ген, кодирующий белок зубцов «застежки-молнии» млекопитающих (SCP1), внедрить в соматические клетки, растущие на искусственной питательной среде, и активировать его, то внутри культивируемых клеток возникает мощная сеть из белков SCP1, «застегнутых» между собой так же, как в центральном пространстве комплекса. Формирование слоя из сплошных белковых «застежек-молний» в культуре клеток означает, что предсказанная нами способность белков комплекса к самосборке доказана [4].
В 1989 и в 2001 гг. сотрудники нашей лаборатории О. Л. Коломиец и Ю. С. Федотова исследовали естественный «демонтаж» синаптонемных комплексов на завершающих этапах их существования. Этот многоэтапный процесс лучше всего удалось проследить на материнских клетках пыльцы в пыльниках ржи, где есть частичная синхронность мейоза [5]. Выяснилось, что латеральные элементы комплекса демонтируются путем постепенного «раскручивания» белковой суперспирали, имеющей три уровня упаковки (рис. 6).
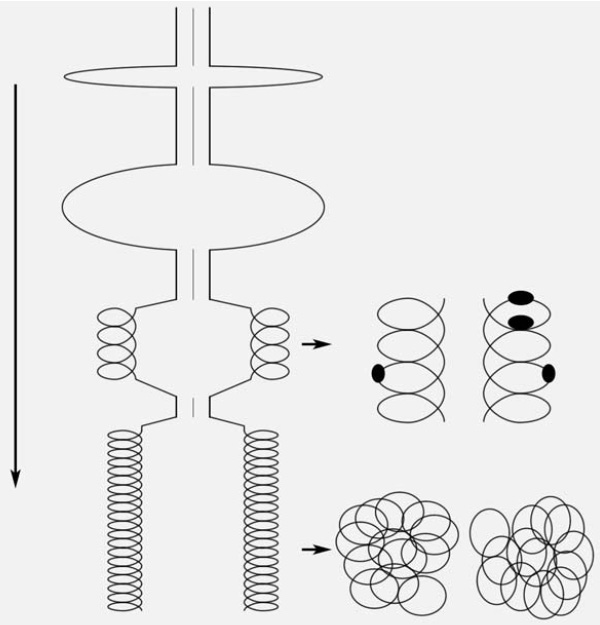 Рис. 6.Этапы демонтажа синаптонемного комплекса в ходе стадии диплотены мейоза у ржи. Все линии обозначают только постепенно вытягивающуюся белковую нить. Изображение: «Природа»
Рис. 6.Этапы демонтажа синаптонемного комплекса в ходе стадии диплотены мейоза у ржи. Все линии обозначают только постепенно вытягивающуюся белковую нить. Изображение: «Природа»
|
|
Основа протяженных латеральных элементов — комплекс из четырех белков когезинов (от англ. cohesion — сцепление). Накануне мейоза в хромосомах появляется специфичный белок когезин Rec8, который заменяет соматический когезин Rad21. Затем к нему присоединяются три других белка-когезина, присутствующие и в соматических клетках, но вместо соматического когезина SMC1 появляется специфический для мейоза белок SMC1b (его N-конец на 50% отличается от N-конца соматического белка SMC1). Этот когезиновый комплекс располагается внутри хромосомы между двумя сестринскими хроматидами, удерживая их вместе. С комплексом когезинов связываются мейоз-специфичные белки, которые становятся мажорными белками хромосомных осей и превращают их (эти оси) в латеральные элементы синаптонемного комплекса [6]. У млекопитающих мажорные белки синаптонемного комплекса — SCP2 и SCP3, у дрожжей белки Hop1 и Red1, а мейоз-специфичный белок — Rec8.




