Генетичні основи нейрогуморальної регуляції статевої поведінки комах.
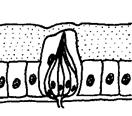
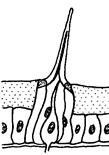
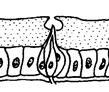
Для більшості комах статева поведінка зв'язана, насамперед, з пошуком статевого партнера. За наявності відповідного гормонального тла рухова активність тварин на етапі такого пошуку різко підсилюється. Але локомоторні реакції, що супроводжують пошук статевого партнера, наближення до нього, відалення від нього й інші, є звичайними руховими реакціями. Вибір шлюбного партнера — одне із самих відповідальних завдань у житті будь-якої комахи, здатної до такого вибору. Системи комунікації, що дозволяють тварині вибирати оптимальних партнерів, бувають досить складними. Вони можуть включати багато різних типів сигналів: · звукові (пісні) · зорові (шлюбне розфарбовування) · поведінкові (спеціальні пози, танці) · хеморецепторні (різні запахи, феромони, а також специфічні молекули, що несуть персональну інформацію). При цьому зміст кожного сигналу може залежати від контексту, тобто від інших сигналів, що вкрай утрудняє для дослідників їхню розшифровку. У хімічній комунікації комах важливу роль грають вуглеводневі феромони. Наприклад, Drosophila melanogaster продукує десятки різних вуглеводнів, що різняться довжиною вуглецевого ланцюга, його розгалуженістю, числом і положенням подвійних зв'язків. Всі вони, вочевидь, беруть участь у хімічному спілкуванні мух, однак про значення конкретних молекул поки відомо небагато. Було з'ясовано, що одні вуглеводні виробляються переважно самками, інші — самцями. «Чоловічий» вуглеводень 7-T (Z-7-Трикозен) приваблює самиць, але відлякує самців. «Жіночі» вуглеводні 7,11-HD (7Z,11 Z-Гептакозадієн) і 7,11-ND (7Z,11 Z-Нонакозадієн), вочевидь, приваблюють самців. Генетично модифіковані самці, що виділяють такі жіночі феромони, сприймаються нормальними самцями як самиці й зазнають сексуальних домагань. Було встановлено, що самці під час парування передають самицям порцію феромону cVA (cis -vaccenyl acetate), що відлякує інших самців і підвищує ймовірність того, що дана самка «збереже вірність своєму першому партнерові». Основну допомогу в пошуках партнера багатьом комахам здійснює нюх. Сприймають запахи комахи за допомогою плакоїдних, трихоїдних, базиконічних і целоконічних сенсил. (Рис. 5.15., 5.16., 5.17). Сенсили (від лат. sensilis - що відчуває) - найпростіші шкірні органи чуття у безхребетних, що виконують роль органів дотику, смаку, нюху. Всі сенсили виникли з одного первісного типу, шляхом спеціалізації для сприйняття певного виду подразнення. Сенсила являє собою видозмінену ділянку покривів тіла (волосок, ямка, пластинка й т.п.), до якого підходять відростки одного або декількох чутливих нейронів. Зовнішня частина утворена клітинами покривів і представлена волоском (тактильні сенсили) або тонкою кутикулярною пластинкою, часто у формі куполу або конусу, зануреною в спеціальну ямку (нюхові й смакові сенсили).
Рис. 5.15. Плакоїдні нюхові сенсили
Рис. 5.16. Трихоїдні нюхові сенсили
Рис. 5.17. Целоконічні нюхові сенсили
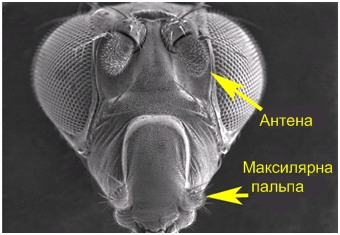
У деяких видів комах для сприйняття запахів служать особливі нитки, що оперізують, чутливі конуси, кутикулярні вирости й ще більш складні утворення. Всі ці дистантні хеморецептори розташовуються майже винятково на антенах. Так, наприклад, рецепторна частина нюхової системи імаго дрозофіли включає дві пари головних придатків (Рис. 5.18). Близько 1200 ольфакторно-чутливих (ольфакторний від лат. Olfactorius - нюховий) нейронів локалізовані на кожному із третіх антенальних сегментів, причому вони асоційовані в кластери по 2-4 нейрони, морфологічно відособлені як ольфакторні сенсили (волоски).
Рис.5.18. Органи нюху дрозофіли
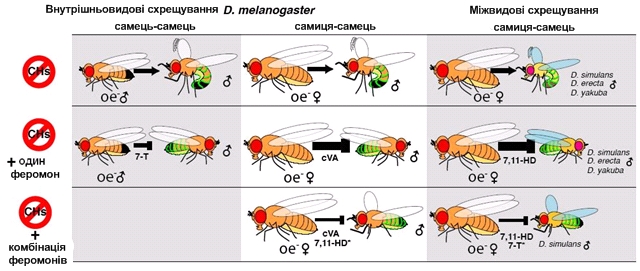
Активація ольфакторно чутливих нейронів відбувається при взаємодії феромону або іншої летючої речовини з одним з ольфакторних рецепторних білків (OR), що експресується на поверхні дендрита. Від кожного ольфакторно-чутливого нейрона відходить роздвоєний аксон, гілки якого кінчаються у відповідних зонах двох антеннальних часток головного мозку - первинному центрі обробки нюхової інформації. Потім проекційні нейрони проводять нервовий імпульс до латеральних ріжків і грибоподібних тіл, що представляють собою вищий нервовий центр, де й відбувається остаточний аналіз ольфакторної інформації і її трансформація в поведінкову відповідь. Чутливість комах до багатьох запахів того ж порядку, що й у людини, іноді - трохи вище, іноді — нижче. Однак чутливість комах до статевих атрактантів може бути вкрай високою. Наприклад, самець шовковичного шовкопряда здатний розрізняти статевий атрактант самиці свого виду – бомбікол – у концентрації 100 молекул на 1 см3.. Самець грушевої сатурнії здатний розрізняти феромони самки на відстані 5-10 км. Такій надчутливості комах до специфічних запахів відповідає дуже строга диференціація ольфакторних стимулів. Аксони відповідних сенсорних нейронів проводять нервові імпульси до нюхових долей дейтероцеребрума (це другий, або середній, відділ головного мозку, що іннервує антени) безпосередньо, без синаптичних перемикань. Слід зазначити, що статеві феромони Drosophila не є летючими речовинами; вони екскретуються на поверхню кутикули й реєструються особинами протилежної статі вже в процесі виконання ритуалу залицяння за допомогою контактних хеморецепторів, розташованих на ротових органах і лапках. Експерименти з генетично модифікованими мухами Drosophila melanogaster, позбавленими здатності виробляти вуглеводневі феромони, дозволили частково розшифрувати хімічну мову комах, за допомогою якої вони обирають шлюбних партнерів. Одна з головних труднощів у вивченні цієї хімічної мови комах полягає в тому, що експериментаторам украй важко маніпулювати складом вуглеводнів, що перебувають на поверхні кутикули. Щоб створити мух, не здатних виробляти вуглеводневі феромони, учені вставили в геном дрозофіли особливу генетичну конструкцію, що повинна була знищити еноцити — спеціалізовані клітини, що розташовуються під кутикулою черевця в комах. Вважається, що саме еноцити відповідають за синтез вуглеводневих феромонів. Оскільки еноцити є модельним об'єктом, використовуваним для вивчення генетичної регуляції індивідуального розвитку, відомі промотори (регуляторні ділянки генів), що активізуються тільки в еноцитах і ніде більше. Щоб одержати мух без еноцитів, учені вставили в геном дрозофіли ген Wrinkled, активізація якого змушує клітину зробити апоптоз, об'єднаний з «еноцитовим» промотором. У результаті в ході розвитку комахи клітини, що перетворилися на еноцити, незабаром гинуть. Спочатку всі генетично модифіковані комахи гинули на стадії личинки. Імовірно, личинки дрозофіл не можуть жити без еноцитів. Генетичну конструкцію ускладнили так, щоб ген апоптоза включався тільки в дорослих мух, а в личинок залишався неактивним. Після цього вченим удалося одержати живих дорослих дрозофіл, позбавлених еноцитів (oe–). І самці, і самиці oe– виявилися майже повністю позбавлені вуглеводневих феромонів. Тим самим була підтверджена гіпотеза про те, що саме еноцити відповідають за їхнє виробництво. Виключення склав чоловічий феромон cVA, що виробляється не еноцитами, а клітинами статевої системи. Мух oe– використовували для вивчення ролі окремих вуглеводнів. З'ясувалося, що відсутність вуглеводневих феромонів не приводить до видимих змін шлюбної поведінки (залицяння). Самці oe– зовсім нормально залицялися до самиць дикого типу, лише небагато поступаючись в активності «диким» самцям. Правда, особливим успіхом у самиць вони не користувалися: у середньому в самця oe– ішло вчетверо більше часу на те, щоб спокусити самицю, у порівнянні з диким самцем. Таким чином, відсутність феромонів у самця майже не впливає на його власну поведінку, але знижує його привабливість для самиць. Ставши малоцікавими для самок, «дезодоровані» самці oe– при цьому стали досить привабливими для самців (як диких, так і інших oe–). Зважаючи на все, самці oe– сприймаються іншими самцями як самки. Взаємні залицяння самців oe– приводять до вигадливих трансформацій статевої поведінки. Наприклад, два самці можуть довго кружляти, упершись один у одного головами. Самець oe– перестає бути об'єктом гомосексуальних домагань, якщо обробити його чоловічим феромоном 7-T. Це підтверджує роль, що передбачалася раніше, 7-Т у запобіганні гомосексуальних взаємодій у самців дрозофіл. Дезодоровані самиці oe– з погляду диких самців стали зовсім непереборними («сексуальними гіперстимулами»). При наявності вибору самці воліли паруватися з такими самками, а не зі звичайними, що виділяють різноманітні вуглеводні. Це значить, що жіночі вуглеводневі феромони існують не для залучення самців, а навпаки — для їхнього стримування. Можливо, це дозволяє самиці в більш спокійній обстановці розібратися, чи підходить їй цей самець. Імовірно, феромони можуть містити інформацію про статеву й видову приналежність, а їхній вплив на сексуальну привабливість може бути різним залежно від контексту. Щоб перевірити ці припущення, учені обробляли «дезодорованих» мух oe– жіночим феромоном 7,11-HD (який дотепер уважався «жіночим афродизіаком», тобто засобом приваблення самців) і чоловічим феромоном cVA, за допомогою якого самці відлякують від своєї партнерки інших залицяльників. Раніше було показано, що феромон cVA дійсно знижує привабливість самиць, однак дія його не абсолютна: багато самиць у природі все-таки відкладають яйця, запліднені декількома різними самцями. Виявилося, що сам по собі жіночий феромон 7,11-HD не впливає на привабливість самиць oe–. Однак він істотно зменшує відлякувальний ефект феромона cVA. Самиці oe–, оброблені феромоном cVA, не викликали інтересу в самців. Однак якщо самку oe– обробити одночасно обома феромонами — 7,11-HD і cVA — те вона стає такою ж привабливою для самців, як і дикі самиці. Очевидно, сексуальна привабливість самиці визначається сполученням сигналів-репелентів, за допомогою яких самець намагається відвернути від її інших самців, і сигналів, виділюваних самицею, які пригнічують дію репелентів. У цьому проявляється конфлікт інтересів самців і самиць. Самці зацікавлені у вірності своїх партнерок, а самицям вигідно мати більше широкий вибір генів для свого потомства, тому вони воліють спаритися з декількома самцями. Дослідникам удалося показати роль вуглеводневих феромонів у формуванні міжвидової репродуктивної ізоляції. Для цього схрещували D. melanogaster (диких і oe–) з мухами із трьох близьких видів: D. simulans, D. yakuba і D. erecta. Із цих трьох видів у двох перших взагалі немає специфічних жіночих вуглеводневих феромонів, а в третього виду вони є, але не такі, як в D. melanogaster. Самці всіх трьох видів або зовсім не хотіли затицятися до диких самиць D. melanogaster, або робили це дуже неохоче. Однак вони охоче залицялися до самиць oe–. Отже, вуглеводневі феромони, виділювані самицями D. melanogaster, беруть участь у підтримці репродуктивної ізоляції між видами: вони відлякують «чужих» самців.
Рис.5.18.а. Роль вуглеводневих феромонів у формуванні міжвидової репродуктивної ізоляції. Верхній ряд: позбавлені еноцитів і кутикулярних вуглеводнів самці й самиці є «сексуальними гіперстимулами» для диких самців D. melanogaster, а також для самців трьох близьких видів. Середній ряд: обробка одиночними феромонами знижує привабливість мух oe–. Нижній ряд: додавання другого феромону пригнічує ефект першого, і в результаті привабливість мух знову зростає.
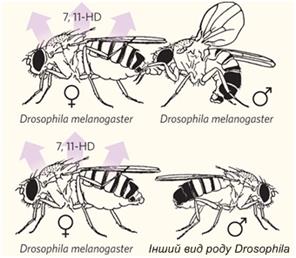
Функцію «відлякування» може виконувати жіночий феромон 7,11-HD. Якщо самицю oe– обробити цим феромоном, вона перестає приваблювати самців чужих видів. Більше того, якщо тим самим феромоном обробити самицю D. simulans, D. yakuba або D. erecta, вона стає непривабливою навіть для своїх рідних самців. Але самиця кожного із чотирьох видів, оброблена цим феромоном, стає бажаним партнером для самців D. melanogaster. Самці D. melanogaster виявляють цікавість до самиць D. erecta навіть без попередньої обробки феромоном 7,11-HD. Можливо, це пояснюється тим, що самиці D. erecta, як і самиці D. melanogaster, продукують інший специфічний жіночий феромон — 7,11-ND. Таким чином, феромон 7,11-HD інформує самців не тільки про статеву, але й про видову приналежність самиць D. melanogaster.
Рис.5.19. Феромон 7,11-HD інформує самців про видову приналежність самиць D. melanogaster. Самиці D. melanogaster виділяють вуглеводневий феромон - речовину 7,11-HD (7, 11-гептакозадієн), що відлякує самців інших видів, але не «своїх».
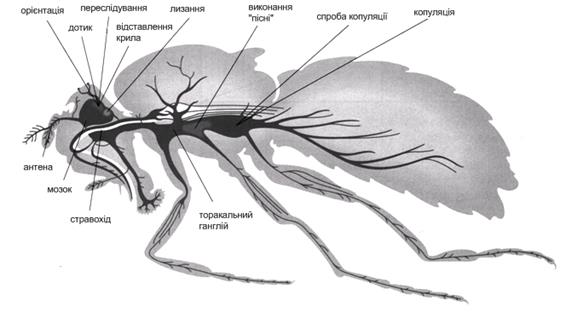
У різних видів дрозофіл ті самі вуглеводневі феромони можуть виконувати різні функції. Наприклад, феромон T-7 у D. melanogaster виробляється переважно самцями, приваблює самиць і відлякує інших самців. У D. simulans цей феромон виробляється більшою мірою самицями й не відлякує, а приваблює самців. Виявилося, що якщо намазати самицю oe–феромоном T-7, це робить її лише небагато більш привабливою для самців D. simulans. Якщо таку ж самицю намазати феромоном 7,11-HD, то це робить її нецікавою для самців всіх видів, крім «свого». Однак феромон T-7 має здатність різко зменшувати відразливу дію феромона 7,11-HD на самців D. simulans. Іншими словами, ефективність феромона T-7 залежить від того, чи була самиця oe– попередньо оброблена феромоном 7,11-HD: якщо так, то феромон Т-7 різко підвищить її привабливість для самців D. simulans, якщо ні — ефект буде набагато слабший. Очевидно, що можливість складних взаємодій між феромональними сигналами характерна не тільки для D. melanogaster. Імовірно, ця можливість закладена в загальній архітектурі системи комунікації за допомогою вуглеводневих феромонів та відповідних рецепторів. Логіка феромональної комунікації в дрозофіл заснована на тім, що всі мухи спершу є сексуально привабливими одне для одного (принаймні для самців), і на цю загальну вихідну привабливість феромони накладають «додаткову» інформацію про статеву й видову приналежність. При цьому зміст окремих сигналів може залежати від контексту, що включає безліч інших сигналів: хімічних, зорових, звукових і інших. Як відбувається передача феромонального сигналу? Припускають наявність двох основних молекулярних компонентів -феромон-звязуючого білка (PBP – pheromone binding protein) і продукту одного з генів Or (Olfactory receptor) сімейства, - за допомогою яких відбувається передача феромонального сигналу. Наприклад, у якості феромон-звязуючого білка, який опосередковує дію такого агрегаційного феромону дрозофіли як цис-вакценіл ацетат (cis-vaccenyl acetate), відомий LASH - білок. Імаго дрозофіли з функціонально неактивним геном lash демонструють знижену агрегаційну активність. Більш того, з електрофізіологічних записів lash - мутантів установлено, що нейрони трихоїдних сенсил, які у нормі експресують LASH, у цих мутантів не стимулюються під впливом цис-вакценіл ацетату. Штучне ж введення LASH у трихоїдну сенсилу таких комах призводить до повного відновлення цис-вакценіл ацетат опосередкованих електричних відповідей. Ці дані свідчать про необхідність даного білка для активації ольфакторно чутливих нейронів, що експресують специфічний рецептор до комплексу цис-вакценил ацетат/LASH. Аграгаційна й харчова поведінка являють собою дуже схожі поведінкові реакції - рух у напрямку до бажаного джерела запаху, але біологічне значення кожної із цих реакцій унікально. Для самців і віргіних (незапліднених) самиць агрегаційна поведінка - це спосіб підвищення шансів спаритися. Для запліднених самиць агрегаційні феромони - це сигнал місць яйцекладки, використовуваних іншими самицями. Завдяки такому явищу як агрегаційна поведінка імаго для личинок, що виходять із яєць, створюються оптимальні умови росту. Тобто створюється оптимальна щільність популяції, що важливо для якості їжі, низького рівня смертності й нормальної швидкості розвитку. Розпізнавання агрегаційного феромону повинне контролюватися тими ж самими центральними мозковими структурами, які активуються при розпізнаванні харчових атрактантів. Імовірно наступні поведінкові реакції (яйцекладка, спарювання) повинні ініціюватися іншими феромонами, оскільки мухи досягли джерела агрегаційного феромона, тобто, виявлення джерела агрегаційних феромонів є завершальним актом відповідної пошукової поведінки. Більш вагому допомогу в пошуках статевого партнера багатьом комахам здійснює зір. Своєрідне забарвлення ряду метеликів, нічне світіння жуків-світляків, тощо - все це, в більшості випадків, ознаки й сигнали для впізнання й приваблення особин свого виду. Комахи, які мають добре розвинені органами слуху, нерідко видають для залучення статевого партнера ті або інші звуки. Це - цвіркуни, коники, цикади й багато хто ін. У певні періоди статевого циклу всі основні органи чуттів комах служать одній меті: знайти особину протилежної статі свого виду. Всі сигнали для зустрічі особин протилежної статі характеризуються, як правило, високою видовою специфічністю, а система виявлення цих сигналів - разючою завадостійкістю. Цікавим є нейро-гормональний контроль ритуалу залицяння. Залицяння самців являє собою комплекс уроджених поведінкових актів, що залучає багато сенсорних модальностей: зорову, контактну хеморецепцію й звукову рецепцію. Послідовність стадій репертуару залицяння, а, отже, і порядок залучення тих або інших сенсорних систем не є випадковою. Загалом схема нейрологічного контролю репертуару залицяння D. melanogaster представлена на Рис.5.20.
Рис. 5.20. Схема нейрологічного контролю окремих стадій репертуару залицяння дрозофіли.
На першій стадії залицяння - стадії орієнтації - ключовою сенсорною модальністю є зорова. Основними органами зору в комах на стадії імаго служать складні очі, додатковими - дорсальні (тім'яні) вічка. Структурною й функціональною одиницею фасеткового ока комах є оматидій (від грецьк. omma, род. відм. - ommatos - око). Оматидії різняться в різних рядів комах. Поля зору в мух перекриваються лише на рівні зорових центрів (у першому оптичному ганглії - у ламіні). Такі очі одержали назву нейросуперпозиційних. Імовірно, таким шляхом у мух досягається підвищення абсолютної чутливості ока без збільшення розміру фасеток і зниження гостроти зору. Фоторецептори комах передають сигнали до зорових центрів без допомоги нервових імпульсів, электротонічно. Більшість дослідників вважають, що сигналом у фоторецепторах слугують рецепторні потенціали, що мають достатню амплітуду й здатні електротонічно градуально активувати зорові інтернейрони. Перетворення зорової інформації починається вже на рівні фоторецепторів, кожний з яких відбирає для передачі у вищі центри лише певну ознака стимулу. Крім того, у деяких комах між фоторецепторами одного оматидію можуть спостерігатися електричні зв'язки, що забезпечують сумацію потенціалів певних рецепторів і сприяють тим найкращому виділенню корисного сигналу із загального набору сигналів.
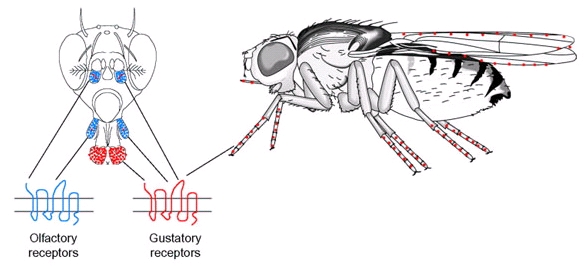
Подальша й більш глибока обробка зорової інформації здійснюється на рівні оптичних гангліїв. Сигнали від фоторецепторів, що завершуються в I оптичному ганглії, надходять на вхід уніполярних нейронів I порядку. Вважається, що аксони фоторецепторів у місці синаптичних перемикань виділяють якийсь гальмівний медіатор, оскільки в уніполярних нейронах у відповідь на засвітлення ока виникають гіперполяризаційні потенціали. Як і у фоторецепторах, потенціали дії в інтернейронах I порядку не генеруються, і, таким чином, ці нейрони електротонічно передають сигнали до інших інтернейронів зорових центрів. На уніполярних інтернейронах конвергють сигнали від декількох фоторецепторів, у результаті чого тут відбувається сумація цих сигналів, і світлочутливість інтернейронів I порядку виявляється вищою в порівнянні з рецепторами. Важливу роль у первинній обробці інформації на рівні I оптичного ганглія грає й латеральне гальмування, що найбільш виражено між сусідніми нейроматидіями. «Геометрія» гальмівних латеральних зв'язків специфічна в різних видів комах, що дозволяє їм найбільше ефективно виділяти в зображенні значимі саме для них ознаки. Відомості про локальні ознаки об'єкта, що надаються нейроматидіями, підлягають подальшій обробці й аналізу. За рахунок горизонтальних (тангенціальних) інтернейронів ці відомості підсумовуються, причому є інтернейрони, що утворюють синаптичні зв'язки із різною кількістю нейроматидіїв і навіть із усіма нероматидіями ганглія. Функціональні завдання, розв'язувані такими підсумовуючими інтернейронами II і більш високого порядку, можуть бути найрізноманітнішими. Установлено, що в зоровій системі комах є нейрони-детектори, здатні виділяти із усього обсягу зорової інформації лише певні ознаки стимулів або сполучень таких ознак. Нейрони-детектори знайдені не тільки в оптичних гангліях, але й у середній частині переднього мозку. Наприклад, виявлені нейрони-детектори руху, високочастотні детектори руху, нейрони-детектори форми. Крім цих узкоспеціалізованих зорових інтернейронів у комах виявлені інтернейрони «загальної» дії, що реагують на зміни освітлення. У комах виявлені полімодальні інтернейрони, що реагують як на зорову, так і на яку-небудь іншу стимуляцію, а також інтернейрони, що не відносяться безпосередньо до зорової системи, але здатні модулювати чутливість і активність зорових інтернейронів. За допомогою такого роду нейронів зоровий аналізатор комахи включається в цілісну систему регуляції поведінки тварини. Перехід до наступної стадії репертуару залицяння, тобто генерація рухового акту відповідно до отриманої сенсорної інформації, очевидно, здійснюється за участю комплексу центрального тіла. Саме туди приходять аксони із всіляких частин мозку, і ця структура забезпечує безпосередні зв'язки з підглотковим ганглієм і гангліями тулубного мозку. Репертуар залицяння D. melanogaster включає дві феромон-опосередковані стадії: стадія дотику й стадія лизання, причому в цьому випадку феромони не є летючими речовинами. Тому, для їхнього розпізнавання необхідний тісний фізичний контакт або, принаймні, максимальне наближення джерела феромонів на черевці самиці й рецепторного нейрона, локалізованого на нозі самця, якщо мова йде про стадію дотику, або на лабелярних пальпах, у випадку стадії лизання. Феромони D. melanogaster синтезуються в еноцитах і потім транспортуються у епідермальні клітини кутикули, звідки секретуються на її поверхню. Специфічний для самців феромон під час парування переноситься на поверхню черевця самиці, і, вочевидь, функціонує як один з інгібіторів залицяння стосовно неї з боку інших самців. Гормональна регуляція синтезу феромонів у комах здійснюється переважно за допомогою трьох основних сполук: ювенільного гормону, 20-гідроксіекдизону й нейрогормону пептидної природи – PBAN (pheromone biosynthesis activating neuropeptide). У D. melanogaster гормональна регуляція продукції феромонів вивчена не дуже докладно, імовірно в ній беруть участь ювенільний гормон, 20-гідроксіекдизон і якийсь поки не встановлений поки мозковий фактор. Сприйняття статевих атрактантів самцями Drosophila відбувається за допомогою контактних хеморецепторів (смакових рецепторів). Первинним смаковим органом є лабелум (губа) – пара пальп у дистальній частині пробосциса (хоботка). Пальпа містить 31 смакову сенсилу - щетинки, - кожна з яких складається з одного механо - і 2-4 хемосенсорних нейронів, оточених допоміжними клітинами. Смакові сенсили підрозділяють на різні морфологічні класи: S (small) - малі; I (intermediate) - середні; L (large) - великі. Додаткові хемочутливі клітини, виявлені на медіальній поверхні кожної пальпи й на внутрішній поверхні глотки, очевидно, вони більшою мірою мають відношення до харчування й контактують безпосередньо з їжею при її поглинанні. У Drosophila смакові щетинки є також на лапках і по краю крила. Саме вони мають безпосереднє відношення до процесу залицяння, кількість смакових щетинок на передніх лапках у самців значно вище, ніж у самиць. У самців – 50, а в самиць - 37. Крім того, нейрони, асоційовані із цими щетинками експресують специфічний для самців gustatory receptor (Gr) ген, продукт якого функціонує як рецептор феромонов. Смакові рецепторні нейрони (подібні ольфакторно чутливим нейронам) - це сенсорні нейрони I типу, об'єднані в смакові сенсили й експресують смакові рецептори - GRs (gustatory receptors). Ці рецептори специфічно взаємодіють із розчинними речовинами, наприклад, цукрами, амінокислотами й іншими. Всі смакові рецепторні нейрони лабіальних пальп і деякі, локалізовані на лапках, проектують аксони у підглотковий ганглій мозку - первинний центр обробки смакової інформації, але аксони деяких смакових рецепторних нейронів, локалізованих на лапках, закінчуються в грудному ганглії.
Рис. 5.21. Переважна локалізація нюхових і смакових сенсил у дрозофіли
Ще одна стадія репертуару залицяння, про регуляцію якої варто згадати – стадія вібрації. У Drosophila відсутні спеціалізовані звуковідтворюючі органи. Звуки ж, видавані самцями дрозофили на цій стадії, є побічним наслідком роботи м'язів крила. Керування системою видавання звуків з боку нервової системи полягає в запуску цих м'язів і підтримці їхньої роботи в певному ритмі. Роботою м'язів крила комах безпосередньо управляють ганглії грудного відділу нервового ланцюжка. Однак рецептивні поля, що беруть участь у активації нейронів цих м'язів, перебувають на значному видаленні від гангліїв, на певних ділянках тіла комахи. На великій кількості видів комах (бабках, тарганах, сарані, осах, бджолах, метеликах, мухах, жуках) було показано, що саме кінцівки є тією інстанцією, що опосередковує запуск і припинення літальних рухів. Очевидно, система ініціації польоту у дрозофіли адаптована до виконання залицяння. Сегментарні ганглії комах містять всі необхідні механізми, що забезпечують навіть дуже складні форми поведінки, але ця сегментарна координаційна система не здатна здійснювати адекватну реакцію комахи на зміни в зовнішнім середовищі без участі вищих центрів. Найбільш важливими центрами регуляції локомоторної активності є грибоподібні тіла й центральне тіло, зв'язані між собою антагоністичними взаєминами, а також підглотковий ганглій. Надглотковий і підглотковий ганглії, не визначаючи здебільшого картини тієї або іншої форми м'язової активності, регулюють її рівень за допомогою гальмівних або збудливих неспецифічних впливів. Іншими словами, вищі локомоторні центри модулюють типи рухів, обумовлені сегментарними гангліями, забезпечуючи тим самим поведінку комахи як єдиного організму.
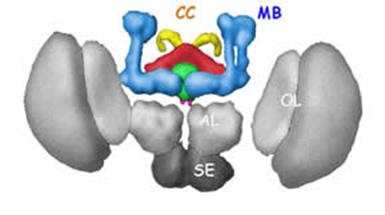
Рис. 5.22. Схематичне тривимірне зображення головного мозку дрозофили: MB (mushroom bodies) - грибоподібні тіла. CC (central complex) - центральний комплекс, що включає чотири елементи: червоним позначені віялоподібні твільця; зеленим – еліпсоподібні тільця; рожевим - noduli; жовтим – протоцеребральний міст. AL (antennal lobes) – антенальні долі. SE (subesophageal ganglia) – підглотковий ганглій; OL (optic lobes) – оптичні долі.
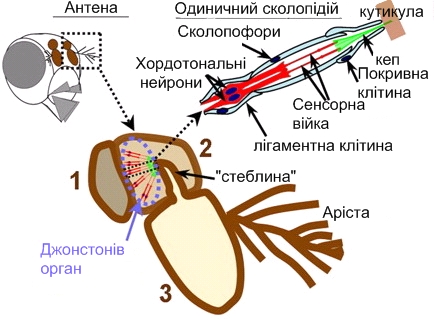
Сприйняття звуків, зокрема, у двокрилих здійснюється за допомогою джонстонових органів, які являють собою найбільш складно влаштовані хордотональні органи й локалізуються в другому членику антен. За своєю будовою, ці органи подібні в різних видів комах. До складу кожної сенсили джонстонова органу входять 3 сенсорних нейрони й 3 обслуговуючі клітини; дендрити нейронів несуть на вершинах по одній видозміненій війці, покритій зовні кутикулярною оболонкою. Вершина цієї оболонки прикріплюється до спвчленовної мембрани, що з'єднує другий і третій членики антени. Фонорецептори джонстонова органу реагують не на абсолютну величину звукового тиску, як у хребетних тварин, а на його зміни - градієнт тиску. Імпульси збудження передаються нейронами джонстонова органу безпосередньо до головного мозку, де й перебувають кілька вищих центрів аналізу слухової інформації. Очевидно, слухові центри функціонально пов'язані й з грибоподібними тілами, оскільки перехід до наступної стадії залицяння - стадії лизання - супроводжується зниженням рухливості самиць, а саме грибоподібні тіла є основним джерелом гальмівних впливів на локомоторну активність комах.
Рис.5.23. Схематичне зображення розташування й будови джонстонова органу дрозофіли.
У керуванні поведінковими реакціями в комах, так само як і в інших високоорганізованих тварин, існують різні рівні, тобто ієрархія командних систем керування. Подібний поділ функцій усередині центральної нервової системи біологічно обґрунтований, оскільки звільняє головні ганглії (головним чином надглотковий) для виконання найбільш складних функцій, пов'язаних з умовно-рефлекторною діяльністю, пам'яттю й ін. Для комах це особливо важливо, тому що кількість нейронних елементів, включених у інтегративну діяльність, у них відносно невелика. Незважаючи на деяку фрагментарність знань з нейрофізіології внутрішніх органів комах, імовірно, керування ними з боку нервової системи є досить складним, та здійснюється в тісній взаємодії з гуморальними факторами. На відміну від керування локомоторною активністю найбільшого значення тут набуває вегетативна нервова система. Послідовність подій при керуванні статевою поведінкою комах загалом така. Всі зміни в зовнішнім середовищі тварина вловлює за допомогою відповідних рецепторів. Інформація про ці зміни передається в мозок, де вона аналізується, і за допомогою ендокринних органів (прилегаючих і кардіальних тіл) створює необхідне (для даного етапу) гормональне тло, забезпечуючи готовність комахи до здійснення тієї або іншої стадії статевого циклу. Однак для реалізації певних рухових реакцій, що супроводжують статеву поведіфнку, необхідні додаткові відомості про навколишнє оточення, наприклад, про наявність статевого партнера й т.п. Ці відомості комаха також одержує за допомогою органів чуттів. Сигнал передається до мозку, який запускає відповідні реакції, надаючи їм, якщо потрібно, того або іншого гормональне забарвлення. Таким чином, безпосередня регуляція статевої поведінки комах забезпечується складним комплексом нейрогуморальних впливів. Питання для повторення й обговорення 1. Як здійснюється вплив нервової системи на функціонування залоз внутрішньої секреції у комах? 2. Що ви знаєте про генетичні основи нейрогуморальної регуляції статевої поведінки комах? 3. Як здійснюється хімічна комунікація комах? 4. Яку роль в поведінці комах грають статеві феромони? 5. Чи можуть феромони брати участь у формуванні міжвидовий репродуктивної ізоляції? 6. Яка роль феромонной комунікації у дрозофіл? 7. Як здійснюється нейро-гормональний контроль ритуалу залицяння в D. Melanogaster? 8. Чи існує в управлінні поведінковими реакціями у комах різні рівні, тобто ієрархія командних систем управління?
|