
Характеристика семейства. Семейство Retroviridae
Семейство Retroviridae
Название семейства происходит от латинских слов retros – обратный и virus – вирус, что связано с наличием в составе вирионов ретровирусов фермента обратной транскриптазы. Классификация. Семейство Retroviridae состоит из 7 родов: Alpharetrovirus, Betaretrovirus, Gammaretrovirus, Deltaretrovirus, Epsilonretrovirus, Lentivirus и Spumavirus, включающих вирусы, патогенные для позвоночных, в том числе и человека. Ретровирусы, вызывающие доказанные инфекционные процессы у людей, входят в рода Gammaretrovirus, Deltaretrovirus, Lentivirus и Spumavirus.
Белки капсида несут перекрестно реактивные (группоспецифические) эпитопы. Геном диплоидный: представлен 2 идентичными копиями линейной несегментированной однониточной «+»-РНК длиной 8,0-9,0*103 нуклеотидов. Изолированная РНК не инфекционна. На концах геномной РНК имеются длинные концевые повторы (LTR). В составе РНК находятся общие для всех ретровирусов структурные гены: gag (группоспецифические антигены), pol (ген полимеразы (синоним – обратная транскриптаза)), env (ген суперкапсида) и v-onc (вирусный онкоген). Наряду со структурными генами, имеются функциональные. Геном нестабилен, что лежит в основе изменчивости, вирулентности и резистентности ретровирусов к специфической противовирусной терапии. Представители отдельных родов ретровирусов могут образовывать «квазивиды». Репродукция ретровирусов. Ретровирусы проникают в клетку путем рецепторного эндоцитоза. Начальные этапы репликации происходят в цитоплазме клетки, где обратная транскрипция вирусной «+»-РНК сопровождается образованием «-»-ДНК. С молекул «-»-ДНК при участии ДНК-зависимой ДНК-полимеразы образуется нить «+»-ДНК. «+»- и «-»-нити ДНК соединяются с образованием двойной нити и транспортируются в ядро клетки. Интеграция вирусной ДНК в ДНК клетки-хозяина осуществляется клеточными эндонуклеазой и лигазой. Интегрированная вирусная ДНК транскрибируется клеточной ДНК-зависимой РНК-полимеразой в «+»-РНК. Полноразмерная «+»-РНК может входить в состав дочерних вирионов, а также выполнять функцию иРНК. Транскрипция и трансляция ретровирусной «+»-РНК ведет к образованию структурных белков и ферментов, которые участвуют в сборке вириона. Информационная «+»-РНК может быть подвергнута РНК-спайсингу – удалению интронов, в результате чего образуются малые молекулы информационной РНК. Самосборка вирионов происходит в цитоплазме. Зрелые вирионы выходят из клетки путем почкования от цитоплазматической мембраны. Онкогены ретровирусов. В состав геномов ретровирусов могут входить онкогены, которые подразделяются на группы: 1) ядерные онкогены, 2) факторы роста, 3) протеин-тирозинкиназы, 4) цитокиновые рецепторы, 5) адапторные белки, 6) серин-треонинкиназы. Ядерные онкогены (ErbA, Ets, Fos, Jun, Maf, Myb, Myc, Rel, Qin, Ski) кодируют белки, локализованные в ядре и способные взаимодействовать с ДНК и РНК клетки-хозяина. Онкоген ErbA связан с онкогеном ErbB. Эти 2 онкогена находятся между gag и env и отделены между собой с помощью интронной последовательности гена ErbB. В гене ErbA имеется 2 точечные мутации в домене, отвечающем за синтез ДНК и 11 мутаций в домене, отвечающем за синтез гормонов. Эти изменения ведут к потере способности синтезировать гормоны, но не влияют на синтез ДНК. ErbA сам по себе не является опухолеродным, но при взаимодействии с ErbB способен вызывать эритролейкемии и фибросаркомы у птиц. Онкоген Ets кодирует фактор транскрипции, который участвует в построении участков ДНК. Ets трансформирует клетки, изменяя экспрессию четко регулируемых генов путем встраивания в участках синтеза. Продукты онкогена Jun локализуются в ядре и имеют несколько доменов. Домен активации необходим для трансформации с помощью Jun. Онкогенная активация Jun происходит из-за нарушения 27 аминокислот на N-конце. Этот участок содержит сайт МАР-киназы, контролирующей активность гена путем фосфорилирования. Онкоген Maf встраивается в вирусный геном в области gag и кодирует лейциновый белок, локализующийся в ядре. Онкоген Ski может трансформировать клетки и вызывать дифференцирование миоцитов. Продукты онкогена активируют транскрипцию, что влияет на дифференциацию клеток. Факторы роста (Sis) трансформируют клетки путем аутокринной активации эндогенного рецептора фактора роста. Обнаружены в вирусе саркомы обезьян. Онкогены, кодирующие протеин-тирозинкиназы (Abl, ErbB, Eyk, Fes/fps, Fgr, Fms, Kit, Ros, Src, Sea, Yes), которые локализуются, в основном, на мембранах трансформированных клеток. Abl-онкоген выделен из вируса лейкемии мышей, обнаружен и у кошек. Белки Abl являются активными тирозинкиназами, обладающими трансформирующими свойствами; располагаются в цитоплазматической мембране клеток. In vitro белки Abl могут трансформировать фибробласты, гемопоэтические клетки лимфоидного и миелоидного рядов. Ген Abl может перестраиваться с помощью сбалансированной транслокации между хромосомами 9 и 22 в более чем в 90 % случаев при злокачественном миелолейкозе у человека. ErbB-онкоген первично отвечает за возникновение опухолей. Его онкогенный потенциал связан с точечными мутациями и делециями внутри цитоплазматического участка Еrb-белка. ErbB является укороченной версией рецептора эпидермального фактора роста - гена семейства тирозинкиназы. Для проявления полного онкогенного потенциала необходима киназная активность и мембранная локализация продукта еrbB. Продукт онкогена Fms является рецепторной тирозинкиназой фактора роста, который активируется с помощью макрофаг-колониестимулирующего фактора. Для трансформации необходимы мембранная локализация и активация тирозинкиназы. Онкоген Src расположен между геном env и 3’-концом вирусного генома. Клеточный Srс является цитоплазматической протеиновой тирозинкиназой, которая участвует в онкогенезе. Онкоген Fgr кодирует активность цитоплазматической тирозинкиназы, что ведет к трансформации. In vitro трансформирует большое количество фибробластов млекопитающих. Уровни белков Fgr повышаются в 50 раз при лимфопролиферативных заболеваниях, ассоциированных с вирусом Эпштейна-Барра. Онкоген Fes/fps способен проникать в gag, что ведет к активации тирозинкиназы и последующей трансформации клетки. Онкогены, кодирующие цитокиновые рецепторы (Mpl). Цитокиновые рецепторы не нуждаются в постоянной активности тирозинкиназы, однако при активации рецепторов происходит активация тирозинкиназ. Mpl – трансформирующий онкоген миелопролиферативного вируса мышей. Клеточный mpl является рецептором для цитокинов и тромбопоэтина, и играет роль в росте и дифференцировке клеток в мегакариоциты. Вирусный трансмембранный белок Мpl является измененной формой клеточного рецептора mpl. Онкогены, кодирующие адапторные белки (Cbl, Crk). Адапторные белки позволяют взаимодействовать белкам между собой, и часто служат мишенями для тирозиновых протеинкиназ. Онкоген Cbl обнаруживается в ядре трансформированных клеток; стимулирует продукцию факторов роста. Вирусные Cbl замещают клеточные Cbl, что ведет к трансформации клетки. Сrk – онкоген вируса саркомы птиц. Активирует клеточную тирозинкиназу и регулирует семейство генов src. Онкогены, кодирующие серин-треонинкиназы (Akt, Raf, Ras). Вирусный онкоген Akt имеет 2 гомолога в клетках человека – Akt1 и Akt2., известные как протеинкиназы В. Эти гены кодируют протеинкиназы С - серин-треонинкиназы, которые отвечают за взаимодействие белков между собой. Вирусный белок Аkt всегда располагается в цитоплазматической мембране.
|
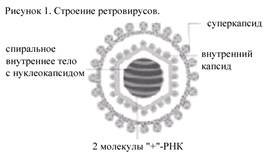 Строение вириона ретровирусов. Вирионы ретровирусов сферической формы, диаметром 80-150 нм, содержат гликолипидный суперкапсид с поверхностными шипами (рис. 1). Гликопротеиновые шипы имеют типоспецифические эпитопы. Внутренняя поверхность суперкапсида выполнена матриксным белком, который называют внешним капсидом. Внутренний капсид может быть сферической, овальной или цилиндрической формы, располагаться центрально или эксцентрично, вероятно, икосаэдрического типа симметрии.
Строение вириона ретровирусов. Вирионы ретровирусов сферической формы, диаметром 80-150 нм, содержат гликолипидный суперкапсид с поверхностными шипами (рис. 1). Гликопротеиновые шипы имеют типоспецифические эпитопы. Внутренняя поверхность суперкапсида выполнена матриксным белком, который называют внешним капсидом. Внутренний капсид может быть сферической, овальной или цилиндрической формы, располагаться центрально или эксцентрично, вероятно, икосаэдрического типа симметрии.


