Функционирование электронотранспортной цепи митохондрий
Для живого организма источником энергии, необходимой для выполнения всех видов работ, является энергия химической связи, высвобождаемая при окислении белков, жиров углеводов. Далее эта энергия используется для синтеза АТФ, а часть рассеивается в виде тепла. Вся биологически доступная энергия из любого органического топлива высвобождается лишь в том случае, если водородные атомы, связанные с углеродом данной молекулы, будут удалены в дыхательную цепь ферментов внутренней мембраны митохондрий, а углерод уйдёт на образование углекислого газа. Все типы окислительно-восстановительных процессов происходят при окислении субстратов в митохондриях, на внутренних мембранах которых размещаются ансамбли из ферментов – дегидрогеназ, коферментов (НАД+, ФАД, УБХ), серии цитохромов b, с1, c и фермента – цитохромоксидазы. Они образуют систему клеточной дыхательной цепи, с помощью которой происходит эстафетная передача протонов и электронов от субстрата к молекулам кислорода, доставленным гемоглобином к клетке. Каждый компонент дыхательной цепи характеризуется определённым значением окислительно-восстановительного потенциала. Движение электронов по дыхательной цепи происходит ступенчато от веществ с низким потенциалом (-0,32 В) к веществам с более высоким потенциалом (+0,82 В), поскольку любое соединение может отдать электроны только соединению с более высоким окислительно-восстановительным потенциалом (таблица 1). Таблица 1 Стандартные редокс-потенциалы биомолекул дыхательной цепи
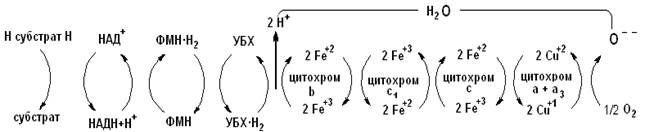
Цепь тканевого дыхания можно представить в виде схемы:
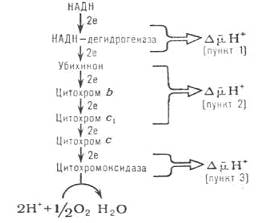
В результате биологического окисления (дегидрирования) два атома водорода (в виде двух протонов и двух электронов) от субстрата поступают в дыхательную цепь. Сначала происходит эстафетная передача протона и пары электронов молекуле НАД+, превращающейся в восстановленную форму НАД × Н, затем системе флавиновых оснований (ФАД/ФАД × Н2 или ФМН/ФМН × Н2), следующим акцептором двух протонов и двух электронов является убихинон (УБХ). Далее происходит передача только электронов: два электрона от УБХ × Н2 принимают на себя последовательно цитохромы в соответствии с величинами их редокс-потенциалов (табл. 1). Последний из компонентов – цитохромоксидаза переносит электроны непосредственно молекуле кислорода. Восстановленный кислород с двумя протонами, полученными от УБХ × Н2 образует молекулу воды. 1/2 О2 + 2 Н+ + 2 е «Н2О Необходимо отметить, что каждая молекула кислорода взаимодействует с двумя электронотранспортными цепями, поскольку в структуре цитохромов возможен только одноэлектронный перенос Fe3+ «Fe2+. Особенностью функционирования дыхательной цепи ферментов является наличие ней участков, где соседние компоненты резко отличаются значениями окислительно-восстановительных потенциалов, именно здесь происходит сопряжение окисления с фосфорилированием АДФ (это комплекс 1 НАДН:убихинон, комплекс 3 убихинол:цитохром с и комплекс 4.цитохромоксидазный комплекс). Такой комплекс, встроенный в мембрану фосфолипида способен функционировать как протонный насос (схема 1). Схема 1. Расположение пунктов сопряжения в цепи дыхательных ферментов.
Согласно теории Митчелла, свободная энергия транспорта электронов в дыхательной цепи затрачивается на перенос из митохондрий через митохондриальную мембрану на ее наружную сторону ионов водорода (протонов). В результате на мембране возникает разность электрических потенциалов Djи разность химических активностей протонов DpH (внутри митохондрий рН выше, чем снаружи). В сумме эти компоненты дают трансмембранную разность электрохимических потенциалов ионов водорода
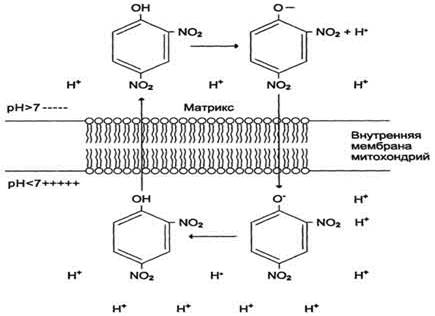
где R -универсальная газовая постоянная, T -абсолютная температура, F- число Фарадея. Величина Перенос электронов (дыхание) Сопряжение тканевого дыхания с окислительным фосфорилированием обеспечивается целостностью внутренней мембраны митохондрий. Это обуславливает возникновение разности потенциалов между её внешней стороной, заряженной положительно, и отрицательно заряженными матриксом и внутренней стороной. Ниже приведена схема трансмембранного переноса протонов и синтеза АТФ в митохондриях.
Дегидрогеназа НАД × Н, расположенная на поверхности мембраны митохондрий, обращенной к матриксу, отдаёт пару электронов на дегидрогеназу ФМН. Это позволяет принять пару протонов из матрикса с образованием ФМН × Н2. Пара протонов, принадлежащих НАД, удаляется на поверхность мембраны. Дегидрогеназа ФМН × Н2 выталкивает пару протонов на цитоплазматическую поверхность мембраны, а пару электронов отдаёт убихинону, который получает возможность присоединить пару протонов из матрикса с образованием УБХ × Н2. УБХ × Н2. выталкивает пару протонов в цитоплазму, а электроны перебрасываются на кислород в матриксе с образованием воды. В результате работы комплекса 1 НАДН:убихинон перекачивается 4 Н+, комплекса 3 убихинол:цитохром с –4 Н+, через комплекс 4 цитохромоксидазный (цитохром а+а3) – ещё 2 Н+. В итоге при переносе двух электронов из матрикса на цитоплазматическую поверхность мембраны перекачивается 10 протонов, что ведёт к созданию разницы потенциалов и разницы рН между поверхностями внутренней мембраны. Разница потенциалов и разница рН обеспечивает движение протонов через протонный канал (фактор F0) в обратном направлении. Движение протонов ведёт к активации АТФ-синтетазы (фактор F1) и синтезу АТФ из АДФ и Н3РО4. Некоторые химические вещества (протонофоры) могут переносить протоны или другие ионы (ионофоры) из межмембранного пространства через мембрану в матрикс, минуя протонные каналы АТФ-синтазы. В результате этого исчезает электрохимический потенциал и прекращается синтез АТФ. Это явление называют разобщением дыхания и фосфорилирования. В результате разобщения количество АТФ снижается, а АДФ увеличивается. В этом случае скорость окисления НАДH и ФАДH2 возрастает, увеличивается количество поглощённого кислорода, но энергия выделяется в виде теплоты. Как правило, разобщители - липофильные вещества, легко проходящие через липидный слой мембраны. Одно из таких веществ - 2,4-динитрофенол, легко переходящий из ионизированной формы в неионизированную, присоединяя протон в межмембранном пространстве и перенося его в матрикс.
Примерами разобщителей могут быть также некоторые лекарства, например дикумарол - антикоагулянт или метаболиты, которые образуются в организме, билирубин - продукт катаболизма тема, тироксин - гормон щитовидной железы. Все эти вещества проявляют разобщающее действие только при их высокой концентрации. Разобщение окислительного фосфорилирования может быть биологически полезным. Оно позволяет генерировать тепло для поддержания температуры тела у новорождённых, у зимнеспящих животных и у всех млекопитающих в процессе адаптации к холоду. У новорождённых, а также зимнеспящих животных существует особая ткань - бурый жир, содержащая много митохондрий. В мембране митохондрий имеется большой избыток дыхательных ферментов по сравнению с АТФ-синтазой. Около 10% всех белков приходится на так называемый разобщающий белок (РБ-1) – термогенин, способный переносить анионы жирных кислот, служащих разобщителями. На внешней стороне мембраны анион жирной кислоты присоединяет протон и в таком виде пересекает мембрану; на внутренней стороне мембраны диссоциирует, отдавая протон в матрикс и тем самым снижает протонный градиент. Разобщение тканевого дыхания и окислительного фосфорилирования наблюдается при охлаждении. Под действием холода стимулируется освобождение норадреналина из окончаний симпатических нервов. В результате происходят активация липазы в жировой ткани и мобилизация жира из жировых депо. Образующиеся свободные жирные кислоты служат не только "топливом", но и важнейшим регулятором разобщения дыхания и фосфорилирования.
|
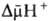 между матриксом митохондрий и внешней водной фазой, разделенных мембраной:
между матриксом митохондрий и внешней водной фазой, разделенных мембраной: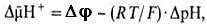
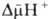 обычно составляет около 0,25 В, причем основная часть (0,15-0,20 В) представлена электрической составляющей Dj. Энергия
обычно составляет около 0,25 В, причем основная часть (0,15-0,20 В) представлена электрической составляющей Dj. Энергия  , выделяющаяся при движении протонов внутрь митохондрий по электрическому полю в сторону меньшей их концентрации, используется АТФ-синтетазой для синтеза АТФ. Таким образом, схему окислительного фосфорилирования, согласно этой концепции, можно представить в следующем виде:
, выделяющаяся при движении протонов внутрь митохондрий по электрическому полю в сторону меньшей их концентрации, используется АТФ-синтетазой для синтеза АТФ. Таким образом, схему окислительного фосфорилирования, согласно этой концепции, можно представить в следующем виде: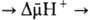 АТФ
АТФ