Работа следящей системы
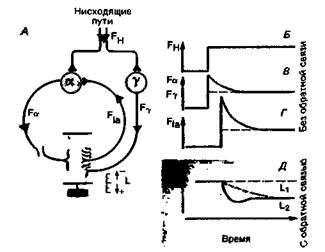
До сих пор мы рассматривали только один аспект действия цепи управления-ее способность поддерживать постоянное значение управляемой переменной. Теперь рассмотрим использование такой цепи в качестве следящей системы. В этом случае, как упоминалось выше, именно изменение эталонного сигнала создает исходное различие между сигналами, поступающими к контроллеру (рис. 15.2, Б). Значение управляемой переменной должно в этом случае измениться на новое заданное. Например, для того чтобы изменился угол сгиба сустава, требуется определенное изменение длины мышцы L. Эталонный сигнал, который может быть использован для программирования меняющегося во времени рабочего значения, представляет собой при этом возбуждение, распространяющееся по нисходящим путям из мозга в мотонейроны в спинном мозгу [13, 17, 18]. Так, процесс регуляции температуры тела, когда последняя повышается при лихорадке, понижается ночью или при гибернации, можно рассматривать как перенастройку заданного значения. Динамика рефлекса растяжения как следящей системы (реакция на ступенчатое воздействие) проиллюстрирована на рис. 15.5. Чтобы последующее обсуждение было понятным, следует увидеть качественное сходство между этой схемой (рис. 15.5, А) и блок-схемами цепей управления на рис. 15.2. Эталонный сигнал передается с помощью двигательных нервных путей, которые идут вниз от головного мозга к спинному (рис. 15.5, А), и может быть представлен в виде изменения частоты разрядов FH в нисходящих аксонах (рис. 15.5, Б). Увеличение или уменьшение активности нисходящих путей вызывает изменение частоты разрядов в а- и γ-мотонейронах в том же направлении (рис. 15.5,5); при этом возникает соответствующее изменение частоты разрядов в афферентных волокнах Iа от мышечных веретен (рис. 15.5,/). Конечный результат такой же, как при внешнем воздействии (см. рис. 15.4),-сокращение или удлинение мышцы (рис. 15.5, Д). Как было отмечено в гл. 5, возбуждение в нисходящих двигательных путях обычно приводит к одновременному синаптическому возбуждению а- и γ-мотонейронов - α-γкоактивации [3, 8]. С точки зрения теории управления оба этих пути передачи супраспинальных команд в систему рефлекса растяжения эквивалентны перенастройке эталон- ГЛАВА 15. ОБЩИЕ ПРИНЦИПЫ РЕГУЛЯЦИИ 339
ного сигнала. Однако система α-γ-коактивации, по-видимому, обладает определенными преимуществами по сравнению с системой, в которой эталонный сигнал заново устанавливается только с помощью контроллера (а-мотонейроны). С одной стороны, интрафузальное сокращение посредством нисходящей активации γ-мотонейронов настраивает диапазон чувствительности или рабочий диапазон мышечного веретена на разные длины мышцы. С другой стороны, поскольку на пропорциональное поведение и на дифференциальное поведение мышечного веретена сложная γ-иннервация оказывает различные влияния, система с коактивацией может быть лучше настроена в отношении оптимальной устойчивости, чем система, в которой эталонный сигнал изменяется только с помощью контроллера. У земноводных нет отдельной γ-системы: у них интрафузальные волокна мышечных веретен и экстрафузальные волокна образуют один общий путь эфферентной иннервации. Наличие отдельной высокодифференцированной γ-двигательной системы следует рассматривать как эволюционное преимущество млекопитающих.
|