
Популяция
ЛЕКЦИЯ Тема: “ПОПУЛЯЦИИ”
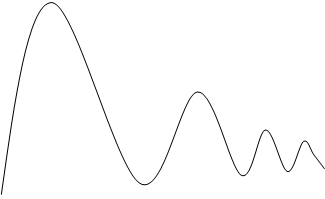
Популяция – любая, способная к самовоспроизведению совокупность особей одного вида, изолированная в пространстве и времени от других аналогичных совокупностей одного и того же вида. Популяция – именно та «ячейка» биоты, которая является основой её существования и элементарной единицей эволюционного процесса, тогда как вид есть его качественный этап. Важнейшими характеристиками являются количественные характеристики популяции – статистические и динамические. Статистические показатели популяции характеризуют состояние популяции на данный момент времени. К ним относятся численность, плотность и показатели структуры популяции. Численность – это число животных или количество растений в пределах некоторой пространственной единицы – ареала, бассейна реки, акватории моря, области и т.д. Плотность – число особей, приходящихся на единицу площади; плотность населения – количество человек, приходящееся на один квадратный километр; плотность гидробионтов – это количество особей в единице объема (в литре или кубометре). Показатели структуры: половой – соотношение полов, размерный – соотношение количества особей различных размеров, возрастной – соотношение количества особей различного возраста в популяции. Численность тех или иных животных определяется различными методами. Например, подсчетом с самолета или вертолета при облётах территории. Численность гидробионтов определяют путём отлавливания их сетями (рыбы), для микроскопических (фитопланктон, зоопланктон) применяют мерные ёмкости. Численность человеческой популяции определяется путём переписи населения. Плотность популяции определяется без учета неравномерности распределения особей на площади или в объеме, т.е. получают среднюю плотность животных, деревьев, населения людей на единицу площади или микроскопических водорослей в единице объема. При уменьшении корма животные расширяют свою территорию. Такое поведение животных называют территориальным поведением. Чем крупнее животное, тем большая ему нужна площадь для добычи пищи, поэтому, чем больше размеры тела особи, тем меньше плотность популяции. Важным условием существования популяции или её экотипа является их толерантность к факторам (условиям) среды. Толерантность у разных особей и к разным частям спектра разная, поэтому толерантность популяции значительно шире, чем у отдельных особей. Свойства популяции значительно отличаются от свойств отдельных особей, что наглядно проявляется в динамике популяций. Динамические показатели популяции характеризуют процессы, протекающие в популяции за какой-то промежуток времени. Основными динамическими показателями является рождаемость, смертность и скорость роста популяции. Рождаемость – это число особей, рождающихся в популяции за единицу времени. При рассмотрении экосистем пользуются другим динамическим показателем – продукцией – суммой прироста массы всех особей из множества популяций биогенного сообщества за определённый промежуток времени. Рождаемость или скорость рождаемости выражают отношением: За бесконечно малый промежуток времени (∆t → 0) получают мгновенную удельную рождаемость b. Размерность этой величины зависит от интенсивности размножения особей: для бактерий – час, для фитопланктона – сутки, для насекомых – неделя или месяц, для крупных млекопитающих – год. Смертность – число особей, погибших в популяции за единицу времени. Смертность – величина обратная рождаемости, но измеряется в тех же величинах и вычисляется по аналогичной формуле: ∆N Но убыль и прибыль организмов зависит не только от рождаемости и смертности, но и от скорости их иммиграции и эмиграции, т. е. от количества особей, прибывших и убывших в популяции в единицу времени. Явления эмиграции и иммиграции на численность влияют несущественно, поэтому ими при расчетах можно пренебречь. Величины рождаемости и смертности по определению могут иметь только положительное значение, либо равное нулю. Скорость изменения численности популяции, т. e. ее чистое увеличение и уменьшение, можно представить и как изменение ∆N за ∆t, а при ∆t → 0 можно ее определить как мгновенную скорость изменения численности, которая может быть рассчитана как: r = b - d. Анализ уравнения показывает, что если b = d, то r = 0, и популяция находится в стационарном состоянии; если же b ≠ d, то r может быть величиной положительной (b ≥ d), и мы имеем численный рост популяции, или отрицательной (b ≤ d), что говорит o снижении численности на данном отрезке времени. Эта формула важна для определения смертности, которую трудно измерить непосредственно, a определить r можно достаточно просто, непосредственными наблюдениями, и d = b - r. Продолжительность жизни вида зависит от условий (факторов) жизни. Различают физиологическую и максимальную продолжительности жизни. Физиологическая продолжительность жизни - это такая продолжительность жизни, которая определяется только физиологическими возможностями организма. Теоретически она возможна, если допустить, что в период всей жизни организма на него не оказывают влияние лимитирующие факторы. Максимальная продолжительность жизни - это такая продолжительность жизни, до которой может дожить лишь малая доля особей в реальных условиях среды. Эта величина варьирует в широких пределах: от нескольких минут y бактерий до нескольких тысячелетий y древесных растений (секвойя), т. e. от 10³ до 10¹¹ сек. Смертность и рождаемость у организмов весьма существенно изменяются c возрастом. Только увязав смертность и рождаемость c возрастной структурой популяции, можно вскрыть механизмы общей смертности и определиться со структурой продолжительности жизни. Такую информацию можно получить c помощью таблиц выживания. Таблицы выживания или «демографические таблицы» содержат сведения o характере распределения смертности по возрастам. Демография изучает размещение, численность, состав и динамику народонаселения, a эти таблицы она использует для определения ожидаемой продолжительности жизни человека. Таблицы выживания бывают динамические и статические. Динамические таблицы строятся по данным прямых наблюдений за жизнью когорты, т. e. большой группы особей, выделяемых в популяции с момента рождения, за короткий промежуток времени относительно общей продолжительности жизни изучаемых организмов, регистрации возраста и до наступления смерти всех членов данной когорты. Такие таблицы требуют длительного наблюдения, измеряемого (для разных животных) месяцами или годами. Но практически невозможно такую таблицу сделать для долго живущих животных или для человека, для этого может потребоваться более 100 лет. Поэтому используют другие таблицы - статические. Статические таблицы выживания составляются по данным наблюдений за относительно короткий промежуток времени за смертностью в отдельных возрастных группах. Зная численность этих групп (сосуществующих когорт), можно рассчитать смертность, специфическую для каждого возраста. Такие таблицы представляют собой как бы временной срез через популяцию. Если в популяции не происходит существенных изменений в смертности и рождаемости, то статические и динамические таблицы совпадают. Данные таблиц выживания позволяют построить кривые выживания. Выделяют три типа основных кривых выживания (рис. 3), к которым в той или иной мере приближаются все известные кривые.
I (дрозофила) человек
II (гидра)
III (устрица) Время
Рис.3. Различные типы кривых выживания.
Кривая I типа, когда на протяжении всей жизни смертность ничтожно мала, резко возрастая в конце жизни, характерна для насекомых, которые обычио гибнут после кладки яиц (ее называют «кривой дрозофилы»), к ней приближаются кривые выживания человека в развитых странах, а также некоторых крупных млекопитающих. Кривая III типа – это случаи массовой гибели особей в начальный период жизни. Гидробионты и некоторые другие организмы, незаботящиеся о потомстве, выживают за счет огромного числа личинок, икринок, семян и т.п. Моллюски, прежде чем закрепиться на дне, проходят личиночную стадию в планктоне, где личинки гибнут в огромных количествах, поэтому кривую III называют еще «кривой устрицы». Кривая II типа (диагональная) характерна для видов, у которыx смертность остается примерно постоянной в течение всей жизни. Такое распределение смертности не столь уж редкое явление среди организмов. Встречаются они среди рыб, пресмыкающихся, птиц, многолетних травянистых растений. Реальные кривые выживания часто представляют собой некоторую комбинацию указанных выше «основных типов». Например, у крупных млекопитающих, у людей, живущих в отсталых странах, кривая I вначале круто падает за счет повышенной смертности сразу после рождения. Еще в XVII в. было установлено, что численность популяций растет по закону геометрической прогрессии, а уже в конце XVIII в. Томас Мальтус (1766-1834) выдвинул свою известную теорию о росте народонаселения в геометрической прогрессии. Эта закономерность роста выражается кривой, изображенной на рис.4. На современном математическом языке эта кривая отражает экспоненциальный рост численности организмов и описывается уравнением:
Nt=Noert, где Nt – численность популяции в момент времени t; No – численность популяции в начальный момент времени to; е – основание натурального логарифма (2,7182); r – показатель, характеризующий темп размножения особей в данной популяции (r-const).
Рис.4. Экспоненциальный рост численности популяции Экспоненциальный рост возможен только тогда, когда r имеет постоянное значение, т.к. скорость роста популяции пропорциональна самой численности:
Таким образом, экспоненциальный рост численности популяции – это рост численности ее особей в неизменяющихся условиях. Условия, сохраняющиеся длительное время постоянными, невозможны в природе. Если бы это было не так, то, например, обычные бактерии могли бы дать такую массу органического вещества, которая могла покрыть весь земной шар слоем толщиной в два метра за два часа. Однако такого в природе не происходит, так как существует множество ограничивающих факторов. Чтобы иметь полную картину динамики численности популяции, а также рассчитать скорость ее роста, необходимо знать величину так называемой чистой скорости воспроизводства (Ro), которая показывает, во сколько раз увеличивается численность популяции за одно поколение, за время его жизни – Т.
Ro=NT/No,
где NT – численность нового поколения; No – численность особей предшествующего поколения; Ro – чистая скорость воспроизводства, показывающая также, сколько вновь родившихся особей приходится на одну особь поколения родителей. Если Ro=1, то популяция стационарная, - численность ее сохраняется постоянной. Скорость роста популяции обратно пропорциональна длительности поколения:
r = lnRo/T,
отсюда ясно, что чем раньше происходит размножение организмов, тем больше скорость роста популяции. Это в равной степени относится и к популяции человека. Воздействие экологических факторов на скорость роста популяции может довести численность популяции до стабильной (r = 0), либо ее уменьшить, т.е. экспоненциальный рост замедляется или останавливается полностью, и J-образная кривая экспоненциального роста как бы останавливается, превращаясь в так называемую S-образную кривую (рис.5) В природе так и происходит: экспоненциальный рост наблюдается достаточно короткое время, после чего ограничивающие факторы его стабилизируют, и дальнейшее развитие популяции идет по логистической модели, что и описывается S-образной или логистической кривой роста популяции. В основе логистической модели (рис.5) лежит простое допущение, что скорость роста популяции (r) линейно снижается по мере роста численности вплоть до нуля при некой численности К. Итак, при начальной численности Nо (близкой к нулю) скорость роста имеет максимальное значение – rmax, а при N = К, r = 0.
rmax
Рис.5. Логистическая модель роста популяции: а - кривая роста численности (N); б – зависимость скорости роста (r) от численности (N).
В результате решения уравнения логистической кривой получаем зависимость:
Nt = K
где Nt – численность популяции в момент времени t; е – основание натурального логарифма; а – постоянная интегрирования. Величину К называют еще емкостью среды в отношении особей данной популяции. Здесь речь идет о биологической емкости среды – степени способности природного или природно-антропогенного окружения обеспечивать нормальную жизнедеятельность (дыхание, питание размножение, отдых и т.п.) определенному числу организмов и их сообществ без заметного нарушения самого окружения (Реймерс, 1990г.). Однако постоянно происходят колебания численности, что отражается в виде колебаний кривой вокруг асимптоты «К» (рис.6), эти колебания
Повышение популяций потенциальной емкости
Потенциальная емкость
Время Рис. 6. Преобразование J-образной кривой роста численности популяции в S-образную кривую при ограничивающем воздействии лимитирующих факторов. называются флуктуациями численности, которые могут быть сезонными и годовыми. Первые обусловлены абиотическими факторами, вторые, плюс к этому, еще и внутренними, биотическими. Колебания, вызванные биотическими факторами, называют осцилляциями. Они отличаются высокой регуляцией и их даже называют циклами. В настоящее время уже достаточно примеров, подтверждающих логистическую модель, как на чисто природных объектах, так и на природно-антропогенных. Например, А. М. Гиляров (1990г.) приводит сведения о размножении северных оленей, интродуцированных (вселенных в местообитания, где они раньше не проживали) на острове Берингова моря. С небольших когорт, состоящих из нескольких десятков особей, в течение ряда лет рост численности по экспоненциальному закону приводил к возникновению популяции оленей, состоящей из нескольких тысяч голов. Затем наблюдалось резкое падение численности тоже до нескольких десятков голов за короткое время, один-три года. Причина – полный расход пищевых ресурсов, которыми обладали эти острова. Экологические стратегии выживания – стремление организмов к выживанию. В 30-х гг. А. Г. Роменский среди растений различал три основных типа стратегий выживания, направленных на повышение вероятности выжить и оставить после себя потомство: виоленты, патиенты и эксплеренты. Виоленты (силовики) – подавляют всех конкурентов; например, деревья, образующие коренные леса. Патиенты – виды, способные выжить в неблагоприятных условиях («тенелюбивые», «солелюбивые» и т.п.). Эксплеренты (наполняющие) – виды, способные появляться там, где нарушены коренные сообщества, - на вырубках и гарях (осины), на отмелях и т.д. Все многообразие экологических стратегий заключено между двумя типами эволюционного отбора, которые обозначаются константами логистического уравнения: r-стратегия и К - стратегия. Тип r-стратегия, или r-отбор, определяется отбором, направленным прежде всего на повышение скорости роста популяции и, следовательно, таких качеств, как высокая плодовитость, ранняя половозрелость, короткий жизненный цикл, способность быстро распространяться на новые местообитания и пережить неблагоприятное время в покоящейся стадии. К - стратегия (или К - отбор) направлена на повышение выживаемости в условиях уже стабилизировавшейся численности. Это отбор на конкурентоспособность, повышение защищенности от хищников и паразитов, повышение вероятности выживаемости каждого потомка, на развитие более совершенных внутривидовых организмов численности. Очевидно, что каждый организм испытывает на себе комбинацию r- и К - отбора, r-отбор преобладает на ранней стадии развития популяции, а К –отбор уже характерен для стабилизированных систем. Логистическая модель роста популяции предполагает наличие некой равновесной (асимптотической) численности и плотности. В этом случае рождаемость и смертность должны быть равны, и если b¹d, то должны действовать факторы, изменяющие либо рождаемость, либо смертность. Факторы, регулирующие плотность популяции, делятся на зависимые и независимые от плотности. Практически, первые – это биотические, а вторые – абиотические факторы. Влияние независимых от плотности факторов хорошо прослеживается на сезонных колебаниях численности планктонных водорослей. Например, в системе Манычских водохранилищ (Северное Предкавказье) диатомовые водоросли дают два «пика» численности – весной и осенью, а в остальное время действуют (точнее, преобладают) зависимые от плотности факторы – конкурентная борьба на выживание с бурно развивающимися летом зелеными и синезелеными водорослями. Помимо выше описанной регуляции существует еще саморегуляция, при которой на численности популяции сказывается изменение качества особей. Различают саморегуляцию фенотипическую и генотипическую. Фенотипы – совокупность всех признаков и свойств организма, сформировавшихся в процессе онтогенеза на основе данного генотипа. При большой скученности (плотности) образуются разные фенотипы за счет того, что в организмах происходят физиологические изменения в результате так называемой стресс – реакции вызываемой неестественно большим скоплением особей. Например, у самок грызунов происходит воспаление надпочечников, что ведет к сокращению рождаемости. Нехватка пищи заставляет особей эмигрировать на новые участки, приводящее к большой их гибели в пути и на новых участках, в новых условиях, т.е. повышается смертность и сокращается численность. Генотипические причины саморегуляции плотности популяции связаны с наличием в ней, по крайней мере, двух разных генотипов, возникших в результате рекомбинации генов. При этом возникают особи, способные размножаться с более раннего возраста и более часто, и особи с поздней половозрелостью и значительно меньшей плодовитостью. Первый генотип менее устойчив к стрессу при высокой плотности и доминирует в период подъема пика численности, а второй – более устойчив к высокой скученности и доминирует в период депрессии. Примером, подтверждающим воздействие генотипических изменений, являются известные с незапамятных времен насекомые – саранча. У саранчовых имеются две разнокачественные группы – одиночная и стадная формы, которые морфологически существенно отличаются. В благоприятные по влажности года преобладают особи одиночной формы, и популяция находится в равновесии. В результате же нескольких подряд засушливых лет создаются условия для развития особей стадной фазы. У стадной формы вылупившиеся из яиц молодые особи (нимфы) быстро двигаются, лучше обеспечены водой и запасами питательных веществ и, хотя у них плодовитость меньше, за счет лучшей выживаемости, более быстрого развития и ярко выраженной способности собираться в группы процесс размножения идет очень быстро и с нарастающей скоростью. Образовавшиеся огромные стаи переносятся ветром на громадные расстояния. Так, мигрирующие очень быстро стаи красной саранчи в Центральной Африке могут занимать площадь, в 1500 раз превышающую области обитания одиночной фазы. Если во время миграции будет найдено место, благоприятное по условиям для размножения, размер стаи может увеличиться до невероятных значений. Так, стая красной саранчи, совершившая налет на Сомали в 1957г., состояла из Циклические колебания можно также объяснить саморегуляцией. Климатические ритмы и связанные с ними изменения в пищевых ресурсах заставляют популяцию вырабатывать какие-то механизмы внутренней регуляции. Так, у мышевидных грызунов Евразии и Северной Америки один период колебаний, состоящий из стадии подъема численности, пика, спада и депрессии длится три-четыре года, иногда пять-шесть лет, а у зайцев – около десяти лет. Одной из известных гипотез такой цикличности является так называемая трофическая (пищевая), утверждающая, что эти циклы зависят не столько от количества пищи, сколько от ее качества. Таким образом, саморегуляция обеспечивается тремя механизмами торможения роста численности: 1) при возрастании плотности и повышенной частоте контактов между особями возникает стрессовое состояние, уменьшающее рождаемость и повышающее смертность; 2) при возрастании плотности усиливается миграция в новые местообитания, краевые зоны, где условия менее благоприятны и повышается смертность; 3) при возрастании плотности происходят изменения генетического состава популяции – замена быстро размножающихся на медленно размножающихся особей. Это свидетельствует о важнейшей роли популяции как в генетико-эволюционном смысле, так и в чисто экологическом смысле.
Популяция лат. populatio население Популяция — это совокупность особей одного биологического вида, способных к свободному скрещиванию и обладающих общим генофондом; П. обладает признаками саморегулирующейся системы и является элементарной единицей эволюционного процесса.
Популяция изогенная — это П., все особи которой генетически идентичны, т. е. гомозиготны по большинству локусов; может возникнуть при длительном инбридинге.
Популяция панмиктическая — это П. раздельнополых организмов, в которой равновероятно формирование любых брачных пар.
Статика популяций Численность. Численностью называют общее число особей в популяции. Существует нижний предел численности, ниже которого популяция не может существовать длительное время. При этом нужно учитывать не всех особей, а только тех, которые принимают участие в размножении – это эффективная численность популяций. Например, если из 100 особей – 50 самцов и 50 самок, то N э. = 100. Если из 100 особей – 90 особей одного пола, а 10 другого, то N э.= 36. Если же из 100 особей на 99 особей одного пола приходится 1 особь другого пола, то N э.= 4. При наличии популяционных волн средняя численность популяции определяется как средняя гармоническая. Обычно численность популяций измеряется сотнями и тысячами особей (такие популяции называют мезопопуляции). У крупных наземных млекопитающих численность популяций может снижаться до нескольких десятков особей (микропопуляции). У растений и беспозвоночных существуют также мегапопуляции, численность которых достигает миллионов особей. У человека минимальная численность популяций составляет около 100 особей. Плотность. В большинстве случаев абсолютную численность популяции определить невозможно. Тогда используют производную характеристику – плотность популяции. Плотность определяется как среднее число особей на единицу площади или объема занимаемого популяцией пространства. В экологии плотность определяется также как масса (биомасса) членов популяции в единице площади или объема. Низкая плотность популяции уменьшает ее шансы на воспроизведение, но увеличивает шансы на выживание. Высокая плотность, наоборот, увеличивает шансы на воспроизведение, но уменьшает шансы на выживание. Следовательно, каждая конкретная популяция должна обладать некоторой оптимальной плотностью. Популяционный ареал Плотность популяции тесно связана с ее пространственной структурой. В популяциях островного типа (с хорошо выраженной границей распространения) плотность распределения особей может быть равномерной. Однако в равнинных популяциях граница распространения всегда размыта. В идеальной популяции можно выделить ее ядро (территория с максимальной плотностью, например, круг), субпериферию (территорию с пониженной плотностью, например, кольцо) и периферию (территорию с низкой плотностью, не обеспечивающей воспроизведение популяции). В реальных популяциях существует множество типов пространственной структуры и, соответственно, типов распределения плотности. Обычно различают следующие типы популяционных ареалов: сплошные, разорванные, сетчатые, кольцевые, ленточные и комбинированные. Динамика популяций Рождаемость. Размножение приводит к появлению в популяции новых особей. Число новых особей, появляющихся в популяции за единицу времени, называется абсолютной рождаемостью. Понятие «новая особь» определяется достаточно произвольно и зависит от видовых особенностей, от целей и задач исследования и других факторов. Например, новой особью (или особью нулевого возраста) может считаться зигота, яйцо, личинка или особь, вышедшая из-под родительской опеки. Отношение числа новых особей к числу имевшихся особей называется относительной (удельной) рождаемостью. Относительная рождаемость может рассчитываться или на одну особь, или на 1000 особей. В ходе размножения численность популяции постоянно изменяется, поэтому вводится понятие мгновенной удельной рождаемости – то есть рождаемости в пересчете на одну особь за бесконечно малый промежуток времени. Этот промежуток зависит от видовых особенностей; для человека достаточно малым промежутком времени считается 1 год. Существуют моноциклические (у растений монокарпические) виды, представители которых размножаются один раз в жизни, и полициклические (у растений поликарпические) виды, представители которых размножаются неоднократно. У раздельнополых диплоидных организмов оценка рождаемости осложняется тем, что для воспроизведения одного потомка требуется пара родителей. В демографии часто учитываются только женские особи. Однако, с точки зрения генетики, самки и самцы в равной степени передают свои гены (аллели) в последующие поколения. Поэтому следует различать плодовитость самок и коэффициент воспроизведения в пересчете на одну особь, независимо от ее пола. Например, в популяции из 500 самцов и 500 самок за единицу времени появилось 1000 особей нулевого возраста. Удельная рождаемость составила одного новорожденного на одну особь, однако каждая самка оставила двух потомков, и каждый самец оставил двух потомков. Численность популяции может увеличиваться не только за счет рождаемости, но и за счет иммиграции особей из других популяций. Существуют зависимые и полузависимые популяции, которые поддерживают и увеличивают свою численность именно за счет иммиграции. Смертность. Смертность – это понятие, противоположное рождаемости. Различают абсолютную смертность (количество погибших особей за единицу времени) и относительную (удельную) смертность (количество погибших особей за единицу времени в расчете на одну особь или на 1000 особей). Характер смертности описывается таблицами и кривыми выживаемости, которые показывают, какая часть новорожденных особей дожила до определенного возраста. Кривые выживаемости обычно строятся в системе координат: «возраст – логарифм числа выживших особей». В этом случае кривые могут быть выпуклыми, вогнутыми и комбинированными. В связи с постоянной смертностью вводится понятие мгновенной удельной смертности, то есть отношению погибших особей к общему числу особей за бесконечно малый промежуток времени (аналогично мгновенной удельной рождаемости). Численность популяции может уменьшаться не только за счет смертности, но и за счет эмиграции особей. Относительный прирост численности. Первоначально при расчете прироста популяции учитывается мгновенная удельная рождаемость и мгновенная удельная смертность (относительные показатели). Тогда прирост популяции называется биотический потенциал, или мальтузианский параметр (r). Для изолированной популяции r = рождаемость – смертность В открытой популяции r = (рождаемость + иммиграция) – (смертность + эмиграция) Прирост популяции может быть положительным, нулевым и отрицательным. Если r > 0, то популяция увеличивает свою численность, если r = 0, то популяция сохраняет стабильную численность, если r < 0, то численность популяции сокращается. Абсолютный прирост численности. Если r величина постоянная (не зависит от численности популяции), то изменение абсолютной численности популяции в единицу времени (dN/dt) и абсолютная численность популяции в данный момент времени (Nt) описываются уравнениями экспоненциального роста. Однако в реальных сообществах всегда существует ограниченность ресурсов. Емкость экологической ниши (К) – это максимально возможная численность популяции в данных условиях. В условиях экологического вакуума (то есть при неограниченности ресурсов среды и при отсутствии конкуренции) величина r остается максимально возможной и постоянной. Но при увеличении численности популяции эта величина снижается; в простейшем случае линейно уменьшается при увеличении численности популяции. В этом случае изменение абсолютной численности популяции описывается уравнением Ферхюльста–Пёрла. Графически эта закономерность отображается логистической (сигмовидной) кривой. Однако в реальных популяциях зависимость r от N и К носит нелинейный характер (эффект группы). Кроме того, при изменении численности происходит изменение экологических характеристик популяции (например, происходит переход с основной пищи на второстепенную), и тогда величина К может измениться. Нужно учитывать также инерционность процессов размножения и гибели, то есть для изменения этих показателей требуется время. За это время может измениться характер действия экологических факторов (например, сезонные или многолетние изменения среды). В природных популяциях могут возникать колебательные процессы (популяционные волны) из-за наличия обратной отрицательной связи между r и N. Уравнение Ферхюльста–Пёрла достаточно точно описывает динамику лишь простых популяций, например, искусственных популяций инфузорий и других мелких организмов с коротким временем генерации в лабораторных условиях. Однако это уравнение помогает выявить основные закономерности роста природных популяций и при введении поправочных коэффициентов достаточно точно прогнозировать их динамику.
Дополнительные факторы, определяющие динамику популяций. На динамику популяции влияют факторы, зависящие и независящие от плотности (численности) популяции. Например, действие климатических факторов в большинстве случаев (но не всегда!) не зависит от плотности популяции. Однако такие факторы как доступность ресурсов, межвидовые взаимоотношения, как правило, зависят от плотности. Популяции видов, у которых рождаемость и смертность в значительной мере зависят от действия внешних факторов, подвержены быстрому изменению биотического потенциала и, соответственно, быстро изменя
|
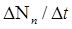 , где
, где  – число особей (яиц, семян и т. п.), родившихся (отложенных, продуцированных и т. п.) за некоторый промежуток времени ∆t. Для сравнения рождаемости в различных популяциях пользуются величиной удельной рождаемости: отношением скорости рождаемости к исходной численности (N):
– число особей (яиц, семян и т. п.), родившихся (отложенных, продуцированных и т. п.) за некоторый промежуток времени ∆t. Для сравнения рождаемости в различных популяциях пользуются величиной удельной рождаемости: отношением скорости рождаемости к исходной численности (N):  .
.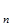 /N∆t, где ∆N
/N∆t, где ∆N 
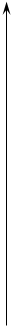 N
N
 60
60 40
40
 0 8 16 24 t
0 8 16 24 t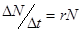 , (r-const.)
, (r-const.)

 К r
К r

 N0 0 Время, t 0 N0 K N
N0 0 Время, t 0 N0 K N (1+e a - rmax·t),
(1+e a - rmax·t), Число особей
Число особей
 «Крах» популяции (или вымирание)
«Крах» популяции (или вымирание)



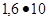 10 особей, и масса ее достигла 50 тыс. т. Если учесть, что за день одна саранча съедает столько, сколько весит сама, то нетрудно представить колоссальные размеры бедствий. Именно такие нашествия насекомых рассматривались как одно из стихийных бедствий на Международном экологическом конгрессе в Йокогаме (1994г.).
10 особей, и масса ее достигла 50 тыс. т. Если учесть, что за день одна саранча съедает столько, сколько весит сама, то нетрудно представить колоссальные размеры бедствий. Именно такие нашествия насекомых рассматривались как одно из стихийных бедствий на Международном экологическом конгрессе в Йокогаме (1994г.).


