ОПИСАНИЕ НЕРВНОЙ СИСТЕМЫ
Головной мозг. У акулы головной мозг состоит из пяти отделов. Передний мозг (telencephalon; рис. 26, 1) крупный, нечетко разделен на правую и левую половины (полушария). В передней части полушарий хорошо заметны сильно развитые обонятельные доли (рис. 26, 2).

Рис. 26. Головной мозг и головные нервы колючей акулы
(со спинной стороны, правый глаз удален):
1 — передний мозг, 2 — обонятельная доля переднего мозга, 3 — промежуточный мозг,
4 — средний мозг, 5 — мозжечок, 6 — продолговатый мозг, 7 — ромбовидная ямка,
8 — спинной мозг, 9 — спинномозговые нервы, 10 — обонятельный мешок,
11 — верхняя косая мышца глаза, 12 — верхняя прямая мышца глаза,
13 — внутренняя прямая мышца глаза, 14 — наружная прямая мышца глаза,
15 — брызгальце, 16 — первая жаберная щель, 17 — вторая — пятая жаберные щели;
I — обонятельный тракт; II — зрительный нерв; III — глазодвигательный нерв; IV — блоковый нерв; VI — отводящий нерв; V — тройничный нерв: г. opht. V — глазничная ветвь, г. max.
V — верхнечелюстная ветвь, г. mand. V — нижнечелюстная ветвь; VII — лицевой нерв: г. opht. VII — глазничная ветвь, г. b. VII — щечная ветвь, r. р. VII — нёбная ветвь, r. hm.
VII — подъязычная ветвь; VIII — слуховой нерв; IX — языкоглоточный нерв;
X — блуждающий нерв; r. branch. X — жаберные ветви, r. spl. X — внутренностная ветвь, r. lat.
X — боковая ветвь
Задняя часть переднего мозга незаметно переходит в промежуточный мозг (diencephalon; рис. 26, 3), который на препарате виден как углубление между вздутиями переднего и среднего мозга.
Средний мозг (mesencephalon; рис. 26, 4) имеет вид крупных парных вздутий (зрительные доли), которые прикрывают собой остальную массу этого отдела мозга.
Четвертый отдел мозга — мозжечок (cerebellum, metencephalon; рис. 26, 5) — у акуловых рыб сильно развит и спереди налегает на средний, а сзади — на продолговатый мозг.
Последний отдел головного мозга — продолговатый мозг (myelencephalon; рис. 26, 6). Сверху в центральной части продолговатого мозга отчетливо видна ромбовидная ямка (рис. 26, 7), представляющая собой полость этого отдела мозга (четвертый желудочек). Она затянута сосудистой оболочкой, которая при препарировке обычно разрушается.
Головные нервы. Характер отхождения головных нервов у акуловых рыб типичен для всех позвоночных животных. От головного мозга у акуловых рыб отходят десять пар головных (черепных) нервов, каждая пара — симметрично с обеих сторон головного мозга. Головные нервы обычно имеют двойное обозначение: порядковый номер и название.
I. Обонятельный нерв (nervus olphactorius). У акуловых рыб, как и у всех позвоночных, от переднего мозга отходят выросты его лобных долей — обонятельные тракты (рис. 26, I). Они образуют у обонятельных капсул расширения — обонятельные луковицы. Обонятельный нерв представляет комплекс коротких чувствующих волокон, идущих от эпителия обонятельного мешка к обонятельной луковице.
II. Зрительный нерв (n. opticus; рис. 26, II). Фактически это зрительный тракт, образующийся как вырост стенки мозга — он отходит от нижне-боковой поверхности промежуточного мозга и, пройдя через отверстие в стенке глазницы, входит в глазное яблоко, распространяясь по внутренней поверхности сетчатки. Нерв чисто чувствующий.
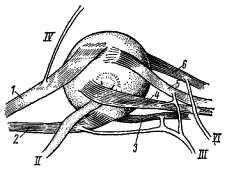
Рис. 27. Глазные мышцы акулы и иннервирующие их нервы:
1 — верхняя косая мышца глаза, 2 — нижняя косая мышца, 3 — нижняя прямая мышца глаза,
4 — внутренняя прямая мышца, 5 — верхняя прямая мышца, 6 — наружная прямая мышца;
II — зрительный нерв, III — глазодвигательный нерв, IV — блоковый нерв,
VI — отводящий нерв
У всех позвоночных животных движения глазного яблока осуществляются при помощи 6 глазных мышц, каждая из которых одним концом прикрепляется к стенке глазницы, а другим — к определенному участку поверхности глазного яблока. От передней стенки глазницы отходят 2 косые мышцы глаза: верхняя (m. obliquus superior; рис. 27, 1) и нижняя (m. obliquus inferior; рис. 27, 2). От задней стенки глазницы пучком отходят 4 прямые мышцы глаза: прямая нижняя (m. rectus inferior; рис. 27, 3), прямая внутренняя (m. rectus internus; рис. 27, 4), прямая верхняя (m. rectus superior; рис. 27, 5) и прямая наружная (т. rectus externus; рис. 27, 6). Эти мышцы иннервируются тремя парами головных нервов — III, IV и VI. Все эти нервы чисто двигательные.
III. Глазодвигательный нерв (n. oculornotorius; рис. 26, ///). Отходит от дна среднего мозга (его видно, если слегка отодвинуть средний мозг от стенки черепа), пронизывает стенку черепа и делится у основания прямых мышц глаза на ветви, идущие к четырем мышцам: к нижней косой и к нижней, внутренней и верхней прямым (рис. 27, ///).
IV. Блоковый нерв (n. trochlearis; рис. 26, IV). В виде тонкой нити отходит от задне-верхней части среднего мозга (на препарате он виден выходящим из-под мозжечка), проходит вперед по крыше среднего моста и, пройдя через переднюю стенку глазницы, разветвляется в верхней косой мышце глаза (рис. 27, IV).
Все остальные головные нервы отходят от продолговатого мозга.
VI. Отводящий нерв (n. abducens; рис. 26, VI). От дна продолговатого мозга сразу уходит в дно черепной коробки. На препарате отхождение этого нерва не видно. Он иннервирует наружную прямую мышцу глаза, на внутренней поверхности которой хорошо видны его ветви (рис. 27, VI).
V. Тройничный нерв (n. trigeminus; pис. 26, V). Это нерв сложный. Он отходит толстым корнем от переднебоковой поверхности продолговатого мозга и сразу же делится на три ветви: глазничную, верхнечелюстную и нижнечелюстную.
Глазничная ветвь (ramus ophthalmicus; рис. 26, r. opht. V) вместе с глазничной ветвью лицевого нерва (см. ниже — VII) проходит через глазницу и ветвится в передней части рыла, иннервируя кожные органы чувств.
Верхнечелюстная (r. maxillaris; рис. 26, г. max. V) и нижнечелюстная (r. mandibularis; рис. 26, r. mand. V) ветви начинаются общим стволом и обособляются позади глаза; они иннервируют мышцы челюстной дуги, зубы, слизистую оболочку ротовой полости и т.д.
По функции тройничный нерв смешанный: он имеет чисто чувствующую ветвь — глазничную и смешанные (чувствующие и двигательные) — верхнечелюстную и нижнечелюстную.
VII. Лицевой нерв (n. facialis; рис. 26, VII). Отходит сразу же за тройничным (на препарате их основания трудно разграничить). Этот нерв также распадается на несколько ветвей. Наиболее крупные и них — глазничная, щечная, нёбная и подъязычная ветви.
Глазничная ветвь (r. ophtalmicus; рис. 26, r. opht. VII) идет вместе с одноименной ветвью тройничного нерва.
Позади глазничной ветви отходит крупный ствол — щечная ветвь (r. buccalis; рис. 26, r. b. VII). Она в виде широкого тяжа проходит по дну глазницы, иннервируя кожные органы чувств брюшной поверхности головы.
Далее общим стволом отходят две ветви: нёбная и подъязычная.
Нёбная ветвь (r. palatinus; рис. 26, r. р. VII) сразу же после обособления уходит в дно черепной коробки и затем ветвится в слизистой оболочке ротовой полости.
Подъязычная ветвь (r. hyomandibularis; рис. 26, r. hm. VII) проходит позади брызгальца (рис. 26, 15) и иннервирует мускулатуру подъязычной дуги и кожные органы чувств боков головы.
Лицевой нерв, как и тройничный, смешанный: его глазничная, щечная и нёбная ветви чувствующие, а подъязычная — смешанная (чувствующая и двигательная).
VIII. Слуховой (n. acusticus; рис. 26, VIII). Идет от боковой поверхности продолговатого мозга и почти сразу же входит в стенку черепной коробки, иннервируя внутреннее ухо. Это чисто чувствующий нерв.
IX. Языкоглоточный нерв (n. glossopharyngeus; рис. 26, IX). Подходит к первой жаберной щели и здесь делится на две ветви, иннервирующие соответственно ее переднюю и заднюю поверхности. Задняя ветвь иннервирует всю мускулатуру первой жаберной дуги. Этот нерв по своей функции смешанный: он имеет и чувствующие, и двигательные волокна.
X. Блуждающий нерв (n. vagus, рис. 26, X). Отходит от задне-боковой поверхности продолговатого мозга несколькими корешками, почти сразу же сливающимися в толстый нервный тяж. В отличие от других головных нервов иннервирует обширную область тела, его основные ветви — четыре жаберные, внутренностная и боковая.
Четыре жаберные ветви (r. branchialis; рис. 26, r. branch. X) иннервируют жаберные щели от второй до пятой. Каждая ветвь, подойдя к жаберной щели, делится на две: передне- и заднежаберную. Переднежаберная ветвь чувствующая, а заднежаберная — смешанная и иннервирует всю мускулатуру соответствующей жаберной дуги (от второй до пятой).
Внутренностная ветвь (r. splanchnicus; рис. 26, r. spl. X) идет как продолжение основного ствола блуждающего нерва. Она входит в брюшную полость, иннервируя внутренние органы.
Более поверхностно лежит боковая ветвь (r. lateralis; рис. 26, r. lat. X), иннервирующая органы боковой линии туловища и хвоста.
Блуждающий нерв смешанный (включает и чувствующие, и двигательные волокна).
У высших позвоночных животных обособляются еще две пары головных нервов.
XI. Добавочный нерв (n. accessorius) имеется только у млекопитающих.
XII. Подъязычный нерв (n. hypoglossus) впервые четко обособляется у пресмыкающихся.
Зачатки этих нервов есть у акуловых рыб в виде нервных корешков, отходящих позади блуждающего нерва в области затылочной стенки мозгового черепа.
Спинномозговые нервы (n. spinalis; рис. 26, 9) видны на препарате — они посегментно отходят от спинного мозга (medulla spinalis). Иннервируют соответствующие сегменты тела и по своей функции представляют собой смешанные нервы, состоящие из чувствующих и двигательных волокон.
Внутреннее ухо (стато-акустический орган). У хрящевых рыб состоит из перепончатого лабиринта, заключенного в хрящевую слуховую капсулу — боковое расширение хрящевого черепа позади глазницы. Три изогнутые трубочки — полукружные каналы — лежат в трех взаимно перпендикулярных плоскостях (рис. 28) и концами открываются в преддверье, или перепончатый мешок.

Рис. 28. Схема перепончатого лабиринта акулы:
1 — полукружные каналы, 2 — овальный мешочек, 3 — круглый мешочек, 4 — лагена
Он подразделяется на две части: верхнюю — овальный мешочек и более крупную нижнюю — круглый мешочек, от которого отходит небольшой вырост — лагена. Полость перепончатого лабиринта заполнена эндолимфой, в которой взвешены мелкие кристаллики — отоконии. В полости круглого мешочка обычно находятся более крупные известковые образования — отолиты. Окончания слухового нерва подходят к отдельным участкам перепончатого лабиринта, покрытым чувствующим эпителием — слуховым пятнам и слуховым гребням. Перепончатый лабиринт — орган чувства равновесия и орган слуха.
ЗАКЛЮЧЕНИЕ
Хрящевые рыбы — новый этап эволюции позвоночных животных. По сравнению с круглоротыми характерен более высокий уровень организации и более активные формы жизнедеятельности.
Наиболее существенное отличие этого класса (и всех челюстноротых - Gnathostomata) от бесчелюстных (Agnatha) в том, что висцеральный отдел черепа челюстноротых состоит из нескольких пар расчленных подвижных висцеральных дуг: челюстной (хватательная функция), подъязычной (функция подвижного прикрипления челюстной дуги к осевому черепу) и нескольких (у хрящевых рыб пяти) жаберных дуг (функция опоры дыхательного аппарата и обеспечения дыхательных движений). Такое устройство висцерального скелета обеспечивает интенсификацию дыхания и значительно облегчает схватывание добычи, тем самым способствуя повышению активности питания и в конечном результате повышению общего уровня обмена веществ.
У хрящевых рыб имеются и другие прогрессивные по сравнению с круглоротыми особенности строения. Появление парных конечностей способствовало увеличению скорости и особенно устойчивости и маневренности плавания. Замена (хотя и неполная) хорды хрящевыми позвонками привела к усилению опорной функции осевого скелета при сохранении его подвижности (способность к изгибам), что также способствовало активизации передвижения, поскольку основным локомоторным органом становится хвостовой отдел тела. Возрастанию скорости плавания способствует и увеличение размеров хвостового плавника.
Резко возрастают у хрящевых рыб относительные размеры головного мозга, все отделы которого хорошо развиты. Органы чувств (глаза, органы обоняния и слуха, сейсмосенсорные органы боковой линии) развиты значительно лучше, чем у круглоротых. Все это также обеспечивает более активный образ жизни. Черепная коробка хрящевых рыб уже закрывает головной мозг со всех сторон; капсулы органов чувств срастаются со стенками мозговой коробки, образуя цельный прочный осевой череп.
Челюстноротым (в том числе и хрящевым рыбам) свойствен иной, нежели у круглоротых, тип строения органов дыхания. Эктодермальные по происхождению жабры прикрепляются к подвижным жаберным дугам и состоят из большого количества тонких жаберных лепестков. Вода непрерывно омывает поверхность жаберных лепестков, что облегчает насыщение крови кислородом. Более отчетливая дифферен-цировка пищеварительного тракта на отделы (как морфологическая, так и функциональная), увеличение поверхности всасывания благодаря имеющемуся в толстой кишке хорошо развитому спиральному клапану, сильное развитие желез пищеварительной системы (печень, поджелудочная железа), вырабатывающих различные пищеварительные ферменты, — все это способствует повышению скорости и эффективности пищеварения.
Вся совокупность прогрессивных изменений, охватывающих различные системы органов, обеспечивает более высокий уровень обмена веществ, общую активизацию жизнедеятельности хрящевых рыб по сравнению с низшими позвоночными (круглоротыми) и появление в этой группе большего разнообразия жизненных форм.
В подклассе пластиножаберных (Elasmobranchii) по характеру образа жизни можно выделить две группы: акулы и скаты. Акулы — в большинстве подвижные хищники, живут в толще воды и отличаются большой скоростью и маневренностью плавания. Для них характерно обтекаемое торпедообразное тело, мощный хвостовой стебель, окаймленный большим хвостовым плавником, острые, направленные назад «хищные» зубы. Питаются акулы разнообразной подвижной добычей (крупной рыбой, головоногими моллюсками и т. д.), которую они ловят в толще воды. Скаты (большинство видов) ведут придонный образ жизни и питаются малоподвижными донными животными; в связи с этим у них уплощенная форма тела и покровительственная окраска спины. Зубы скатов уплощены и не имеют острых вершин. Комплекс зубных рядов образует своеобразную «терку», хорошо приспособленную для раздавливания раковин моллюсков и панцирей иглокожих и ракообразных.
Однако по сравнению с эволюционно бэлее молодым классом костных рыб многие особенности строения хрящевых рыб выглядят примитивными и ограничивают возможности широкой адаптивной радиации. Массивный хрящевой скелет увеличивает массу тела. Пояса парных плавников лежат в толще мускулатуры и не укреплены на осевом скелете. Все это ограничивает возрастание подвижности. Массивные хрящевые челюсти способны лишь к однообразным простым хватательным движениям. Приспособительные изменения, связанные с различным характером питания акул и скатов, ограничиваются различиями в строении зубов.
По сравнению с костистыми рыбами для хрящевых рыб характерен более низкий уровень обмена веществ, что определяется рядом морфологических и физиологических особенностей: еще довольно простым строением дыхательного аппарата (отсутстви жабе рной крышки), меньшей дифференцировкой и длиной кишечника, меньшей скоростью кровотока и кислородной емкостью крови и т. д. Все это находит свое выражение и в ограниченном распространении хрящевых рыб (только обитатели моря), и в малом разнообразии их жизненных форм при ограниченном числе видов (около 600 против почти 20 тыс. видов костных рыб).
Но в море и сейчас хрящевые рыбы могут считаться вполне процветающей группой, успешно отстаивающей свои жизненные ниши и конкурирующей с костными рыбами. Этому способствуют некоторые своеобразные особенности хрящевых рыб. Прежде всего для них характерна большая живучесть, чем у костных рыб. Она проявляется в способности к длительному голоданию, в возможности переносить очень большие повреждения и ранения, в большей мощи мускулатуры (в эксперименте акула заметно не снижает скорости движения, если к ней привязан дополнительный груз, составляющий до 25% массы ее тела; многие костистые рыбы почти полностью теряют способность двигаться при дополнительной нагрузке в 5—10% от массы тела). Эти особенности в той или иной степени связаны с относительно большим размером головного мозга и большей (по сравнению с костными рыбами) его дифференцировкой (сильное развитие переднего мозга и мозжечка и др.), с повышенной автономностью периферической нервной системы, тем что в мышечных сегментах имеется значительное количество дыхательного пигмента — миоглобина, и другими физиологическими и биохимическими особенностями.
Специфические черты в строении половой системы, обеспечивающие возможность внутреннего оплодотворения, относительно крупные размеры яиц с большим запасом питательных веществ, их прочная рогоподобная скорлупа резко снижают эмбриональную смертность. Еще более повышает эффективность размножения свойственное многим видам хрящевых рыб яйцеживорождение. Эти особенности снижают расход энергии при размножении и повышают выживаемость акуловых рыб в борьбе за существование.
Очень своеобразен у хрящевых рыб механизм осморегуляции. Осмотическое давление в тканях и крови хрящевых рыб примерно равно осмотическому давлению морской воды. Этот изотонизм обеспечивается высоким содержанием мочевины в крови и тканях. При изменениях солености среды меняется уровень мочевины в крови и тканях, приводя осмотическое давление жидкостей тела в соответствие с таковым среды. Концентрация же биологически важных солей в организме остается при этом неизменной. Такой тип осморегуляции свойствен только хрящевым рыбам.
Хрящевые рыбы — наиболее древняя группа рыб, для которых характерны многие примитивные черты организации. Они сохранились до наших дней благодаря выработке своеобразных черт организации, частично описанных выше, помогающих им успешно выдерживать конкуренцию с костными рыбами — эволюционно более молодой и прогрессивной группой, имеющей более высокий сбщий уровень организации. Хрящевые и костные рыбы — это два самостоятельных конвергентных типа развития водных позвоночных животных.
Дополнительная литература
Гуртовой Н. Н., Матвеев Б. С, Дзержинский Ф. Я. Практическая зоотомия позвоночных. Низшие хордовые, бесчелюстные, рыбы. М., 1976.
Никольский Г. В. Частная ихтиология. М., 1971.
Никольский Г. В. Экология рыб. М., 1974.
Суворов Е. К. Основы ихтиологии М., 1948.
Шмальгаузен И. И. Основы сравнительной анатомии позвоночных. М., 1947.
КЛАСС КОСТНЫЕ РЫБЫ
OSTEICHTHYES
ТЕМА 6. ВСКРЫТИЕ КОСТИСТОЙ РЫБЫ
СИСТЕМАТИЧЕСКОЕ ПОЛОЖЕНИЕ ОБЪЕКТА
Подтип Позвоночные, Vertebrata
Класс Костные рыбы, Osteichthyes
Подкласс Лучеперые, Actinopterygii
Группа Костистые рыбы, Teleostei
Представитель — Щука, Esox luceus L.
МАТЕРИАЛ И ОБОРУДОВАНИЕ
На одного-двух студентов необходимы:
1. Щука (свежая или фиксированная).
2. Ванночка.
3. Скальпель.
4. Пинцет.
5. Ножницы.
6. Препарировальные иглы — 2.
7. Булавки — 10—15.
8. Вата гигроскопическая.
9. Марлевые салфетки — 1—2.
ЗАДАНИЕ
Познакомиться с особенностями внешнего вида рыбы. Произвести вскрытие; рассмотреть строение основных систем внутренних органов. Сделать следующие рисунки.
1. Внешний вид щуки.
2. Общее расположение внутренних органов.
3. Мочеполовая система самца и самки.
4. Жабра в поперечном разрезе.
ВНЕШНИЙ ВИД
Форма тела костистых рыб очень разнообразна. У щуки (рис. 29) тело вытянутое и слегка сплющенное с боков. Подразделение его на голову, туловище и хвост нечеткое. По бокам головы видны относительно крупные жаберные крышки (рис. 29, 1). Их задний край служит границей головного и туловищного отделов. Костные жаберные крышки покрыты кожей, края которой сзади выступают, прикрывая общее (одно с каждой стороны) жаберное отверстие.
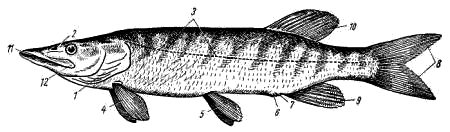
Рис. 29. Внешний вид щуки:
1 — жаберная крышка, 2 — ноздря, 3 — боковая линия, 4 — грудные плавники,
5 — брюшные плавники, 6 — анальное отверстие, 7 — половое и мочевое отверстия,
8 — хвостовой плавник, 9 — анальный плавник, 10— спинной плавник,
11— ротовое отверстие, 12 — свободный задний край верхнечелюстной кости
Во рту щуки находятся острые, загнутые назад зубы. По бокам головы перед глазами располагаются ноздри (рис. 29, 2). Они имеют вид ямок, каждая из которых подразделена поперечной перегородкой на два отверстия. Ноздри не сообщаются с ротовой полостью (проверить введением иголочки или щетинки!).
Туловище щуки покрыто костной чешуей. Каждая чешуйка представляет собой тонкую, округлой формы костную пластинку, которая передним краем укреплена в коже. У щуки свободный (задний) край чешуи гладкий, такая чешуя носит название циклоидной. У некоторых других видов (например, окуня) свободный край чешуи зазубрен, такая чешуя называется ктеноидной. Чешуи снаружи покрыты тонким эпидермисом. При рассмотрении свежего материала хорошо заметно, что тело рыбы покрыто слизью. Слизь выделяется многочисленными одноклеточными кожными железами.
Вдоль боков туловища тянется боковая линия (рис. 29, 3), обнаруживаемая при внешнем осмотре в виде тонких отверстий, прободающих чешуи. Эти отверстия ведут в особые каналы, где располагаются органы боковой линии с нервными окончаниями, воспринимающими колебания окружающей тело воды. На голове боковая линия разделяется на несколько ветвей (надглазничную, подглазничную, подъязычно- челюстную и др.).
По бокам передней части туловища видны парные, относительно небольшие грудные плавники (рис. 29, 4), а ближе к заднему концу туловища — парные брюшные плавники (рис. 29, 5).
На брюшной стороне туловища, каудальнее места прикрепления брюшных плавников, заметно анальное отверстие (рис. 29, 6), а сразу за ним — мочеполовой сосочек; у части рыб он представляет собой углубление (рис. 29, 7) с двумя отдельными отверстиями: мочевым (заднее) и половым. Место расположения заднепроходного, полового и мочевого отверстий служит границей туловищного и хвостового отделов.
Хвостовой отдел имеет непарные плавники: хвостовой (рис. 29, 8) и подхвостовой (рис. 29, 9). У костистых рыб они внешне равны по величине — такой тип хвостового плавника называют гомоцеркальным. Анальный плавник расположен впереди нижней лопасти хвостового плавника. У щуки здесь же, на хвостовом стебле, спереди от верхней лопасти хвостового плавника располагается и непарный спинной плавник (рис. 29, 10). У большинства других костистых рыб этот плавник (иногда их бывает два, три и даже больше) находится на спинной стороне туловищного отдела.
Мышечная система костных рыб в принципе сходна с акуловыми.
ВСКРЫТИЕ
1. Ножницами сделать короткий поперечный разрез брюшной стенки впереди анального отверстия.
2. Осторожно ввести в разрез тупую ветвь ножниц по направлению к голове и, все время прижимая эту ветвь к брюшной стенке (чтобы не повредить внутренностей), сделать разрез вдоль средней линии брюха. Разрез довести до самой передней части так называемого перешейка (часть брюшной стенки, вдающаяся между нижними краями жаберных крышек), перерезав кости плечевого пояса.
3. От начала продольного разреза (у анального отверстия) сделать еще один разрез — вверх до позвоночника.
4. Приподнимая боковую стенку тела, вести разрез вперед вдоль позвоночника до жаберной крышки, отделяя боковую стенку тела.
5. Срезать жаберную крышку.
6. Осторожно с помощью пинцета, скальпеля и иголок освободить препарат от кусков мышц и пленок, мешающих его рассмотреть (особенно аккуратно надо действовать в области сердца и отходящей от него брюшной аорты). Кровь отсасывать ватными тампонами.
7. Последовательно рассмотреть строение различных систем внутренних органов.
ОБЩАЯ ТОПОГРАФИЯ ВНУТРЕННИХ ОРГАНОВ
Кровеносная система. Сердце (cor) располагается в нижней передней части полости тела, в основании перешейка. Венозная кровь собирается в венозной пазухе, или венозном синусе (sinus venosus; Рис. 30, 1; рис. 31, 1). Отсюда кровь переходит в предсердие (atrium; рис. 30, 2; рис. 31, 2) и затем в более толстостенный желудочек (ventri-culus; рис. 30, 3; рис. 31, 5).
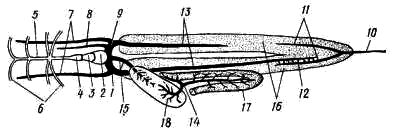
Рис. 30. Схема кровеносной системы костистой рыбы
(вид снизу; не показаны выносящие жаберные артерии,
их слияние в спинную аорту и ветвление последней):
1 — венозная пазуха, 2 — предсердие, 3 — желудочек, 4 — луковица аорты, 5 — брюшная аорта,
6 — приносящие жаберные артерии, 7 — передние кардинальные вены, 8 — яремная вена,
9 — кювьеров проток, 10 — хвостовая вена, 11 — воротные вены почек,
12 — анастомозы между воротной веной правой почки и правой задней кардинальной веной,
13 — задние кардинальные вены, 14 — воротная вена печени, 15 — печеночная вена,
16 — почки, 17 — кишечник, 18 — печень
В отличие от хрящевых костистые рыбы не имеют артериального конуса. Непосредственно от желудочка отходит крупная брюшная аорта (aorta ventralis; рис. 30, 5; рис. 31, 4), образующая в этом месте расширение—луковицу аорты (bulbus aortae; рис. 30, 4). Брюшная аорта отдает четыре пары приносящих жаберных артерий (arteria branchialis afferentia; рис. 30, 6; рис. 31, 6).
При обычной препарировке периферическую часть кровеносной системы, описание которой дается далее, рассмотреть не удается. В жаберных лепестках каждая приносящая жаберная артерия распадается на систему капилляров. Через их стенки происходит газообмен крови с омывающей жабры водой. Обогащенная кислородом артериальная кровь по системе капилляров собирается в выносящие жаберные артерии (arteria branchialis efferentia), которые на спинной стороне впадают в парные корни спинной аорты. Корни аорты в заднем отделе головы сливаются, образуя непарную спинную аорту (aorta dorsalis); она проходит под позвоночником и отсылает многочисленные артериальные сосуды ко всем участкам тела.
Венозная кровь из хвостового отдела идет по непарной хвостовой вене (vena caudalis; рис. 30, 10), которая разделяется на две воротные вены почек (vena porta renalis; рис. 30, 11), входящие в почки. У костистых рыб в отличие от хрящевых воротная система образуется только в левой почке. Из почек кровь по парным задним кардинальным венам (vena cardinal is posterior; рис. 30, 13) направляется вперед. На уровне сердца задние кардинальные вены сливаются с передними кардинальными венами (vena cardinalis anterior), несущими кровь от головы. В результате слияния задних и передних кардинальных вен образуются парные кювьеровы протоки (ductusCuvieri; рис. 30, 9), впадающие в венозную пазуху. В нее же впадает несущая кровь от нижних частей головы нижняя яремная вена (v. jugularis inferior; рис. 30, 8).

Рис. 31. Общее расположение внутренних органов щуки:
1 — венозный синус, 2 — предсердие, 3—желудочек сердца, 4 — брюшная аорта,
5 — луковица аорты, 6 — приносящие жаберные артерии, 7 — кювьеров проток, 8— жабра,
9 — желудок, 10 — двенадцатиперстная кишка, 11 — тонкая кишка, 12 — прямая кишка,
13 — анальное отверстие, 14 — печень, 15 — желчный пузырь, 16 — желчный проток,
17 — поджелудочная железа, 18 — селезенка, 19 — плавательный пузырь, 20 — почка,
21 — мочеточник, 22 — мочевой пузырь, 23 — мочеполовой сосочек,
24 — мочевое отверстие, 25 — половая железа, 26 — половое отверстие
От кишечника кровь по воротной вене печени (vena porta hepatis; рис. 30, 14) попадает в печень, где эта вена распадается на систему капилляров, т. е. образует воротную систему печени. По выходе из воротной системы печени кровь по короткой печеночной вене (vena hepatica; рис. 30, 15) попадает в венозную пазуху. Боковых вен, свойственных хрящевым рыбам, у костистых рыб нет.
У костистых рыб, как и у хрящевых, один замкнутый круг кровообращения. В сердце рыб находится только венозная кровь. Сокращениями сердца эта кровь направляется в жабры, где происходит освобождение от углекислого газа и насыщение кислородом. Насыщенная кислородом артериальная кровь, выходящая из жаберной системы, по многочисленным артериям направляется к различным органам и тканям тела, где происходит обратный процесс: отдача кислорода из крови в ткани и насыщение крови углекислотой, т. е. превращение крови из артериальной в венозную. По системе вен венозная кровь возвращается в сердце. Понятия «артериальная» и «венозная» кровь определяют качественные отличия в газовом составе крови. Эти понятия не всегда совпадают с названиями кровеносных сосудов. Так, по брюшной аорте (артерии) и по приносящим жаберным артериям движется венозная кровь; независимо от состава крови артериями называют сосуды, по которым кровь идет от сердца, а венами — сосуды, по которым кровь направляется к сердцу.
Дыхательная система. Органами дыхания у костистых рыб служат жабры (рис. 31, 8), имеющие, как и у хрящевых рыб, эктодермальное происхождение. С каждой стороны располагаются четыре полные жабры; у некоторых рыб рудиментарная полужабра находится на внутренней стороне жаберной крышки.
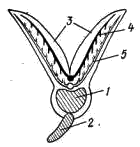
Рис. 32. Схема поперечного разреза жаберной дуги костистой рыбы:
1 — жаберная дуга, 2 — жаберная тычинка, 3 —жаберные лепестки;
4 — приносящая жаберная артерия, 5 — выносящая жаберная артерия
Вырежьте кусок жабры и рассмотрите ее строение. Межжаберных перегородок, характерных для хрящевых рыб, у костных рыб нет. Два ряда жаберных лепестков (рис. 32, 3) своими основаниями прикрепляются непосредственно к костной жаберной дуге (рис. 32, 1) или к рудименту межжаберной перегородки, а их свободные окончания свешиваются в околожаберную полость. Эта полость снаружи прикрыта костной жаберной крышкой, имеющей существенное значение в акте дыхания. С внутренней стороны каждой жаберной дуги имеются многочисленные отростки — жаберные тычинки (рис. 32, 2), идущие по направлению к соседней жаберной дуге. Жаберные тычинки образуют своеобразный цедильный аппарат, препятствующий выходу пищевых частиц из глотки через жаберную полость наружу. У видов, питающихся планктоном (например, сельдей), этот аппарат представлен особенно длинными и густо сидящими тычинками.
Через стенки кровеносных капилляров в жаберных лепестках, как уже говорилось, происходит газообмен крови с омывающей жабры водой. Более крупные сосуды (приносящие и выносящие жаберные артерии) проходит по жаберным дугам в основании жаберных лепестков (рис. 32, 4, 5).
Пищеварительная система. В ротовой полости щуки находятся острые, слегка направленные назад конические зубы. Без четких границ ротовая полость переходит в глотку, прободенную жаберными щелями. В глубине глотки начинается короткий пищевод, который почти сразу же переходит в желудок (gaster; рис. 31, 9). За желудком следует кишечник, слабо дифференцированный на двенадцатиперстную (duodenum; рис. 31, 10), тонкую (intestinum; рис. 31, 11) и прямую (rectum; рис. 31, 12) кишки. Прямая кишка открывается наружу анальным отверстием (рис. 31, 13).
Сразу за сердцем в передней части брюшной полости под желудком располагается крупная печень (hepar; рис. 31, 14). На ее внутренней стороне находится желчный пузырь (vesica lellea; рис. 31, 15) — полость, в которой скапливается вырабатываемая в печени желчь. От желчного пузыря начинается желчный проток (ductus choledochus; рис. 31, 16), впадающий в начало двенадцатиперстной кишки. Вдоль желчного протока лежит поджелудочная железа (pancreas; рис. 31, 17). В месте перехода желудка в двенадцатиперстную кишки (первый изгиб кишечника) рядом с желудком расположена компактная селезенка (lien; рис. 31, 18).
Над кишечником в верхней части брюшной полости находится крупный плавательный пузырь (vesica natatoria; рис. 31, 19), который служит гидростатическим органом. Плавательный пузырь щуки узким протоком связан с передней частью кишечника. У многих других рыб (например, карповых, окуня и др.) во взрослом состоянии плавательный пузырь полностью изолирован от кишечника.
Мочеполовая система. В верхней части брюшной полости по бокам плавательного пузыря располагаются парные половые железы. У самок половые железы представлены длинными яичниками (ovarium; рис. 33, 2), которые имеют хорошо заметную «зернистую» структуру. Задние, вытянутые отделы яичников играют роль выводных протоков (рис. 33, 3) и открываются непарным половым отверстием (рис. 33, 5) позади анального отверстия.

Рис. 33. Задний участок мочеполовой системы самки щуки:
1 — плавательный пузырь, 2 — яичник, 3 — выводной проток яичника,
4 — мочеполовой сосочек, 5 — половое отверстие. 6 — почки, 7 — мочеточник,
8 — мочевой пузырь, 9 — мочевое отверстие, 10 — кишечник, 11 — анальное отверстие
Половые железы самцов — длинные, гладкие, дов086; льно плотные семенники (testis); они занимают такое же положение, как и яичники.
Задние отделы семенников превратились в короткие выносящие протоки, открывающиеся общим половым отверстием позади анального отверстия.
Чтобы рассмотреть почки, нужно удалить кишечник и плавательный пузырь. Почки (ren; см. рис. 31, 20; рис. 33, 6) располагаются на спинной стороне полости тела по обе стороны от позвоночника. По их краю проходят мочеточники (ureter; рис. 33, 7), которые, выйдя из почек, сливаются в единый непарный мочевой проток. Мочевой пузырь (vesica urinaria; рис. 31, 22; рис. 33, 8) представляет собой вырост передней стенки начальной части этого протока. Непарный мочевой проток открывается наружу мочевым отверстием (рис. 33, 9) позади полового отверстия.
ТЕМА 7. СКЕЛЕТ КОСТИСТОЙ РЫБЫ
СИСТЕМАТИЧЕСКОЕ ПОЛОЖЕНИЕ ОБЪЕКТА
Подтип Позвоночные, Vertebrata
Класс Костные рыбы, Osteichthyes
Подкласс Лучеперые, Actinopterygii
Группа Костистые рыбы, Teleostei
Представитель — Судак, Lucioperca lucioperca L.
МАТЕРИАЛ И ОБОРУДОВАНИЕ
Смонтированный скелет судака на подставке — один на всю группу. На одного-двух студентов необходимы:
1. Смонтированный на картонных планшетах разборный скелет судака.
2. Препарировальные иглы — 2.
ЗАДАНИЕ
Рассмотреть скелет в целом.




