http://www.rpi.edu/dept/bcbp/molbiochem/MBWeb/mb2/part1/aacarbon.htm#glucket
Contents of this page:
Glucogenic & ketogenic amino acids
Catabolism of amino acids to:
3-C pyruvate; 4-C & 5-C Krebs cycle intermediates
Other examples: Aromatic amino acids; Methionine
Cofactors discussed include: coenzyme B12, THF, tetrahydrobiopterin, SAM
Amino acids, when deaminated, yield a-keto acids that,directly or via additional reactions, feed into the major metabolic pathways (e.g., Krebs Cycle). Fig. 26-11, p. 995, depicts the ultimate products of degradation of amino acid carbon skeletons. Pathways for all amino acids will not be discussed in detail here. Emphasis will be placed on reactions involving cofactors that are not discussed elsewhere. Amino acids are grouped into two classes, based on whether or not their carbon skeletons can be converted to glucose:
Glucogenic amino acids: Their carbon skeletons are degraded to pyruvate, or to one of the 4- or 5-carbon intermediates of Krebs Cycle that are precursors for gluconeogenesis. Glucogenic amino acids are the major carbon source for gluconeogenesis when glucose levels are low.They can also be catabolized for energy or converted to glycogen or fatty acids for energy storage.
Ketogenic amino acids: Their carbon skeletons are degraded to acetyl-CoA or acetoacetate. Acetyl CoA, and its precursor acetoacetate, cannot yield net production of oxaloacetate, the precursor for the gluconeogenesis pathway. For every 2-C acetyl residue entering Krebs Cycle, two carbon atoms leave as CO2. (For review, see notes on Krebs Cycle.) Carbon skeletons of ketogenic amino acids can be catabolized for energy in Krebs Cycle, orconverted to ketone bodies or fatty acids. They cannot be converted to glucose.
| The amino acids may be grouped for discussion on the basis of the specific keto acid products of their deamination.
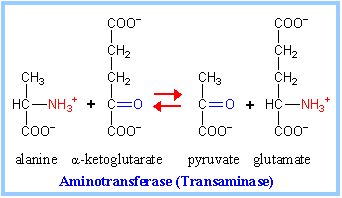
The 3-carbon a-keto acidpyruvate is produced from alanine, cysteine, glycine, serine, & threonine (p. 996).
Alanine deamination via Transaminase directly yields pyruvate (at right).
| 
|
| Serine is deaminated to form pyruvate via Serine Dehydratase.
Glycine, which is also a product of threonine catabolism, is converted to serine by a reaction involving tetrahydrofolate (to be discussed below).
| 
|
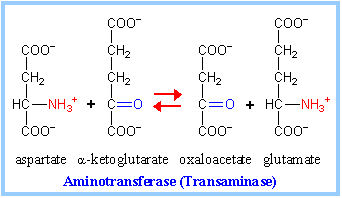
| The 4-carbon Krebs Cycle intermediate oxaloacetate is produced from aspartate & asparagine (p. 1000).
Aspartate deamination via Transaminase directly yields oxaloacetate (at right). Aspartate also is converted to fumarate in the Urea Cycle (p. 992, & notes on amino acid catabolism - N). Fumarate is then converted in Krebs Cycle to oxaloacetate.
| 
|
| Asparagine loses the amino group from its R-group by hydrolysis catalyzed by Asparaginase. This yields aspartate, which can then be converted to oxaloacetate by transamination (above).
| 
|
The 4-carbon Krebs Cycle intermediate succinyl-CoA is produced from isoleucine, valine, & methionine (p. 1002, 1004). Propionyl-CoA, which is an intermediate on these pathways, is also a product of b-oxidation of fatty acids with an odd number of C atoms.
| The conversion of propionyl-CoA to succinyl-CoA is summarized at right.
Propionyl-CoA is carboxylated to yield methylmalonyl-CoA.
A racemase yields the L-isomer essential to the subsequent reaction.
| 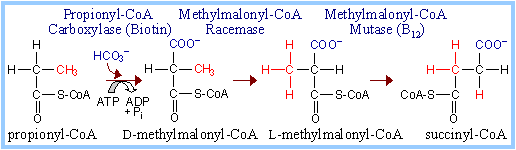
|
| Methylmalonyl-CoA Mutase catalyzes a molecular rearrangement in which the branched carbon chain of methylmalonyl-CoA is converted to the linear carbon chain of succinyl-CoA. The carboxyl that is in ester linkage to the thiol of coenzyme A is shifted to an adjacent carbon atom, with opposite shift of a hydrogen atom.
Recall that coenzyme A is a large molecule, as shown at right.
Coenzyme B12, a derivative of vitamin B12 (also called cobalamin), is the prosthetic group of Methylmalonyl-CoA Mutase.
| 
|
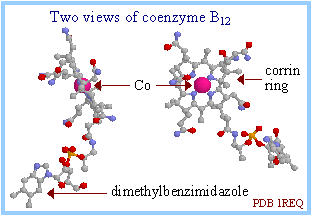
A crystal structure of the enzyme-bound coenzyme B12 is shown at right in ball and stick display, with the cobalt atom in spacefill display. See also diagrams and more information in the Stadtman website at NIH, in a University of Bristol website, and in the Voet & Voet textbook pages 922-927.
Coenzyme B12 contains a heme-like corrin ring with a cobalt ion coordinated to 4 ring nitrogen atoms.
Within the Methylmalonyl-CoA Mutase active site, the cobalt atom of coenzyme B12 has two axial ligands:
- the methyl C atom of 5'-deoxyadenosine (not shown here).
- an enzyme histidine N
When vitamin B12 is free in solution, a ring N atom of the dimethylbenzimidazole group (shown extending away from the Co in the crystal structure of the enzyme at right) serves as an axial ligand to the cobalt. When vitamin B12 is enzyme-bound, a histidine side-chain N substitutes for the dimethylbenzimidazole.
|  Color key: C O N P Co Coenzyme B12 within Methylmalonyl-CoA Mutase Color key: C O N P Co Coenzyme B12 within Methylmalonyl-CoA Mutase
|
| Homolytic cleavage of the deoxyadenosyl C-Co bond yields a deoxyadenosyl carbon radical as Co3+ becomes Co2+. Reaction of this with methylmalonyl-CoA generates a radical substrate intermediate and 5'-deoxyadenosine. Following rearrangement of the substrate, the product radical abstracts a hydrogen atom from the methyl group of 5'-deoxyadenosine. This yield succinyl-CoA and the 5'-deoxyadenosyl radical, which reacts with coenzyme B12 to reestablish the deoxyadenosyl C-Co bond.
Explore at right Methylmalonyl-CoA Mutase, with desulfo-coenzyme A (CoA analog lacking the reactive thiol) present at the active site.
Methyl group transfersare also carried out by B12 (cobalamin). Methyl-B12 (methylcobalamin), with a methyl axial ligand substituting for the deoxyadenosyl moiety of coenzyme B12, is an intermediate of such transfers. For example, vitamin B12 is a prosthetic group of the mammalian enzyme that catalyzes methylation of homocysteine to form methionine (discussed below).
|  Methylmalonyl-CoA Mutase Methylmalonyl-CoA Mutase
|
Vitamin B12 is synthesized only by bacteria. Ruminant animals obtain B12 from bacteria in their digestive system. Humans obtain vitamin B12 from meat or dairy products.
Vitamin B12 bound to the protein gastric intrinsic factor is absorbed by cells in the upper part of the human small intestine via receptor-mediated endocytosis. B12 synthesized by bacteria in the large intestine is unavailable. Strict vegetarians eventually become deficient in B12 unless they consume it in pill form.
Vitamin B12 is transported in the blood bound to the protein transcobalamin, which is recognized by a receptor that mediates uptake into body cells.
The branched chain amino acids initially share in part a common pathway (p. 1004). Branched Chain a-Keto Acid Dehydrogenase (BCKDH) is a multi-subunit complex homologous to the Pyruvate Dehydrogenase complex. Genetic deficiency of BCKDH is called Maple Syrup Urine Disease (MSUD), because high concentrations of branched chain keto acids in the urine give it a characteristic odor.
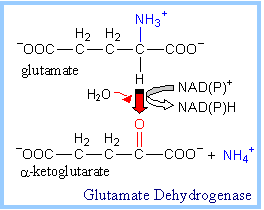
| The 5-carbon Krebs Cycle intermediate a-ketoglutarateis produced from arginine, glutamate, glutamine, histidine, and proline (p. 1001).
Glutamate deamination, via Glutamate Dehydrogenase or Transaminase, directly yields a-ketoglutarate (at right & above).
| 
|
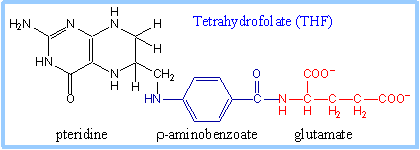
| Histidine is first converted to glutamate. The last step in this pathway involves the cofactor tetrahydrofolate (THF).
THF, which has a pteridine ring, is a reduced form of the B vitamin folate. The conversion of folate to THF is presented on p. 1028.
Within a cell, tetrahydrofolate has an attached chain of several glutamate residues, linked to one another by isopeptide bonds involving the R-group carboxyl.
| 
|
| Tetrahydrofolate exists in various forms in which single-carbon units, of varying oxidation state, are bonded at N5 or N10, or bridging between these nitrogen atoms, as shown on p. 1029 and at right. The cellular pool of THF includes these various forms, that are produced and utilized in different reactions.
In the diagrams at right, N10 with R is r-aminobenzoic acid linked to a chain of glutamate residues (see above).
Reactions that utilize THF as a donor of a single carbon unit include reactions for synthesis of thymidylate, methionine, formylmethionine-tRNA, etc.
| 
|
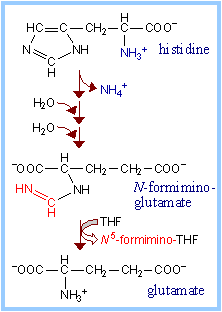
| N 5-formimino-tetrahydrofolate, produced in the pathway for degradation of histidine, is shown at right.
| 
|
| In the pathway of histidine degradation, conversion of N -formiminoglutamate to glutamate involves transfer of the formimino group to tetrahydrofolate (THF), yielding N 5-formimino-THF.
Because of the essential roles of THF as acceptor and donor of single carbon units, dietary deficiency of folate, genetic deficiencies in folate metabolism or transport, and the increased catabolism of folate seen in some disease states, result in various metabolic effects leading to increased risk of developmental defects, cardiovascular disease, and cancer.
| 
|
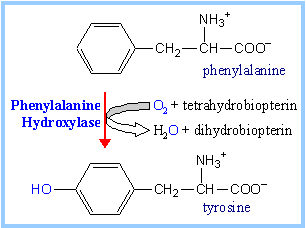
| The aromatic amino acids phenylalanine & tyrosine are catabolized to fumarate & acetoacetate (p. 1009).
Hydroxylation of phenylalanine to form tyrosine involves the reductant tetrahydrobiopterin. Biopterin, like folate, has a pteridine ring. Structures are shown on p. 1010. Dihydrobiopterin is reduced to tetrahydrobiopterin by electron transfer from NADH. Thus NADH is secondarily the electron donor for the conversion of phenylalanine to tyrosine.
Overall the reaction is considered a mixed function oxidation because one O atom of the O2 is reduced to water while the other is incorporated into the amino acid product.
| 
|
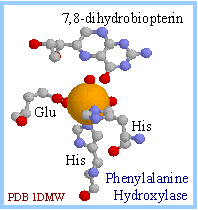
| Phenylalanine Hydroxylase includes a non-heme iron atom at its active site. X-ray crystallographic analysis has shown that the following are ligands to the iron atom (displayed at right in spacefill): His nitrogen, Glu oxygen, and water oxygen atoms (shown in ball & stick).
O2, tetrahydrobiopterin, and the iron atom in the ferrous (Fe++) oxidation state participate in the hydroxylation reaction. O2 is thought to react initially with the tetrahydrobiopterin to form a peroxy intermediate.
| 
|
| Genetic deficiency of Phenylalanine Hydroxylase leads to the disease phenylketonuria. Phenylalanine and phenylpyruvate (the product of deamination of phenylalanine via transaminase) accumulate in the blood and urine. Mental retardation results unless treatment begins immediately after birth.
Treatment consists of limiting phenylalanine consumption to levels barely adequate to support growth. Tyrosine, an essential nutrient for individuals with phenylketonuria, must be supplied in the diet.
Tyrosine is a precursor for synthesis of melanins, and of epinephrine and norepinephrine (structures below). High [phenylalanine] inhibits Tyrosine Hydroxylase, an enzyme on the pathway for synthesis of the pigment melanin from tyrosine. Thus individuals with phenylketonuria have light skin and hair color.
| 
|
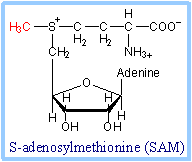
| Methionine is converted to S -Adenosylmethionine (SAM) by an ATP-dependent reaction. SAM serves as a methyl group donor in various synthetic reactions.
The resulting adenosylhomocysteine is hydrolyzed to homocysteine, which may be catabolized via a complex pathway to cysteine and succinyl-CoA.
Or methionine may be regenerated from homocysteine by methyl transfer from N 5-methyl-tetrahydrofolate, via a methyltransferase enzyme that utilizesB12 as prosthetic group. The methyl group is transferred from tetrahydrofolate to B12 to homocysteine.
Another pathway converts homocysteine to glutathione.
| 
|
| In various reactions, S-adenosylmethionine (SAM) is a donor of diverse chemical groups including methylene, amino, ribosyl and aminoalkyl groups, and a source of 5'-deoxyadenosyl radicals. But SAM is best known as a methyl group donor.
| 
|
Examples of the role of S -adenosylmethionine(SAM) as methyl group donor:
- methylation of bases in tRNA
- methylation of cytosine residues in DNA (p. 1205)
- methylation of norepinephrine to form epinephrine (shown at right)
| 
|
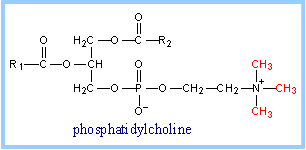
- conversion of the glycerophospholipid phosphatidylethanolamine to phosphatidylcholine (shown at right).
Enzymes involved in formation and utilization of S -adenosylmethionine are particularly active in liver, which has important roles in synthetic pathways involving methylation reactions, and in regulation of blood methionine.
| 
|
In summary, three cofactors act as methyl group donors in synthetic reactions:
- methyl-B12
- S -adenosylmethionine (SAM)
- N 5-methyl-tetrahydrofolate (N 5-methyl-THF)
The complex pathways for degradation of lysine and tryptophan will not be covered.
Studio Exercise:
Explore the National Center for Biotechnology Information's OMIM (Online Mendelian Inheritance in Man) file on the hereditary disease phenylketonuria.
Link: OMIM - PHENYLKETONURIA
Use the menu at the top of the OMIM page to change the display to Clinical Synopsis or Detailed. Within the Detailed display, you may choose to view listed items such as Clinical Features, and Biochemical Features.
Search for answers to the following:
- Which enzyme is deficient in phenylketonuria (PKU1)?
- What is a possible biochemical mechanism of damage to the brain?
- How is phenylketonuria treated and controlled?
- Can treatment be stopped in adult-hood?
- What is the danger with consumption of the artificial sweetener aspartame?
- What is the special risk during pregnancy for mothers with even mild forms of the disease?
- What populations are most at risk?
If you have time search the OMIM database for inborn errors of other enzymes of amino acid catabolism, such as homocystinuria, a defect in an enzyme on the pathway from homocysteine to cysteine.
Линейная механика разрушения
Трещина представляет собой самый острый концентратор напряжений. С уменьшением радиуса кривизны в вершине надреза концентрация напряжений возрастает. Одновременно предполагаем, что материал пока не проявляет свои пластические свойства. Поскольку радиус кривизны вершины трещины весьма мал и не определен, то и напряжения в малой области в непосредственной близости к вершине трещины могут быть большими и неопределенными количественно.
Вместе с тем, если рассматривать напряженную зону вне малой окрестности вершины трещины (диаметром более 10 диаметров зерна для более уверенного применения модели однородного континуума), то на основании решений задач теории упругости оказывается (см. раздел 3.1), что компоненты напряжений 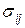 представлены так называемыми асимптотическими формулами в виде
представлены так называемыми асимптотическими формулами в виде
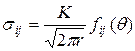 . (3.2.1)
. (3.2.1)
В этих формулах 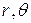 - полярные координаты точки с полюсом в вершине трещины;
- полярные координаты точки с полюсом в вершине трещины; 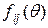 - известные функции, принимающие значение единицы или нуля при
- известные функции, принимающие значение единицы или нуля при  ;
;  - коэффициент интенсивности напряжений, не зависящий от координат
- коэффициент интенсивности напряжений, не зависящий от координат 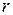 и
и 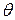 , его размерность: сила/длина в степени
, его размерность: сила/длина в степени  . Перевод К в разные системы:
. Перевод К в разные системы: 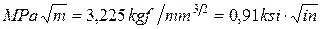 .
.
Из формулы (3.2.1) следует, что при  напряжения
напряжения 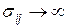 , т.е. напряжения в вершине трещины имеют особенность вида
, т.е. напряжения в вершине трещины имеют особенность вида 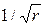 . Коэффициент при этой особенности – коэффициент интенсивности напряжений К – характеризует величину напряжений в целом во всей области, в которой справедливы формулы (3.2.1). Характер же распределения напряжений в этой области в зависимости от и один и тот же для тел любой формы и любой схемы нагружения. Поэтому для характеристики напряженно-деформированного состояния в области справедливости асимптотических формул (3.2.1) вполне достаточно знания коэффициента К; этот коэффициент (как следствие решения линейной теории упругости) прямо пропорционален параметру приложенных к телу нагрузок и зависит от размеров тела, в частности, от размеров трещины. Рост нагрузки приводит к пропорциональному росту К, что в свою очередь означает рост напряжений (рис. 3.9). На нижней эпюре напряжений
. Коэффициент при этой особенности – коэффициент интенсивности напряжений К – характеризует величину напряжений в целом во всей области, в которой справедливы формулы (3.2.1). Характер же распределения напряжений в этой области в зависимости от и один и тот же для тел любой формы и любой схемы нагружения. Поэтому для характеристики напряженно-деформированного состояния в области справедливости асимптотических формул (3.2.1) вполне достаточно знания коэффициента К; этот коэффициент (как следствие решения линейной теории упругости) прямо пропорционален параметру приложенных к телу нагрузок и зависит от размеров тела, в частности, от размеров трещины. Рост нагрузки приводит к пропорциональному росту К, что в свою очередь означает рост напряжений (рис. 3.9). На нижней эпюре напряжений 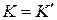 и этот коэффициент одинаков в каждой точке этой кривой. На линии
и этот коэффициент одинаков в каждой точке этой кривой. На линии  коэффициент интенсивности напряжений для каждой точки этой линии один и тот же и равен . На всех точках верхней заштрихованной линии коэффициенты интенсивности напряжений одинаковы и равны вязкости разрушения
коэффициент интенсивности напряжений для каждой точки этой линии один и тот же и равен . На всех точках верхней заштрихованной линии коэффициенты интенсивности напряжений одинаковы и равны вязкости разрушения 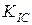 .
.
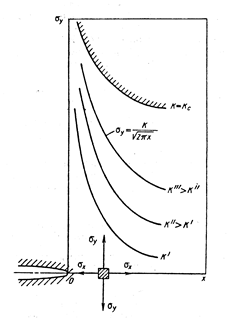
Рис. 3.9. Нормальное напряжение перед вершиной трещины отрыва при разных коэффициентах интенсивности напряжений
Основываясь на этом, Ирвин в 1957 г. предложил силовой критерий разрушения в виде
 . (3.2.2)
. (3.2.2)
Неравенство означает безопасное состояние – отсутствие роста трещины; равенство означает наступление предельного (критического) состояния равновесия, при котором трещина получает возможность распространения.
Слева в выражении (3.2.2) стоит коэффициент интенсивности напряжений К, который следует знать в виде функции нагрузки, размеров детали и трещины, а справа он же, но определенный из опыта и играющий роль механической характеристики материала, оценивающей его трещиностойкость, т.е. сопротивление материала росту в нем трещины[1]. Величина 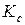 - критический коэффициент интенсивности напряжений для плоского образца данной толщины t (более кратко – “вязкость разрушения”, иногда просто трещиностойкость) – определяется из эксперимента. При хрупком разрушении для трещины отрыва (тип 1) пишут . Методы экспериментального получения статических характеристик трещиностойкости регламентированы в [6].
- критический коэффициент интенсивности напряжений для плоского образца данной толщины t (более кратко – “вязкость разрушения”, иногда просто трещиностойкость) – определяется из эксперимента. При хрупком разрушении для трещины отрыва (тип 1) пишут . Методы экспериментального получения статических характеристик трещиностойкости регламентированы в [6].
Пример. В трубопроводе вдоль образующей обнаружена сквозная трещина длиной 2 l =60 мм. Диаметр трубопровода 1220 мм, толщина стенки 14 мм. Найдем критическое окружное напряжение  и давление p, при котором трещина начнет неуправляемо распространяться, приводя к полному разрушению.
и давление p, при котором трещина начнет неуправляемо распространяться, приводя к полному разрушению.
Проведем мысленный разрез вдоль образующей трубы с противоположной стороны от трещины и развернем трубу до совмещения с плоскостью. Получилась полоса длиной  =3830 мм, а ширину для расчета примем равной b =200 мм. Полоса растягивается напряжением равным окружному в трубе. Материал полосы – сталь 17Г1С, условный предел текучести
=3830 мм, а ширину для расчета примем равной b =200 мм. Полоса растягивается напряжением равным окружному в трубе. Материал полосы – сталь 17Г1С, условный предел текучести 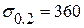 МПам1/2, характеристика трещиностойкости
МПам1/2, характеристика трещиностойкости 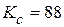 МПам1/2.
МПам1/2.
Из справочника [5] находим формулу для коэффициента К, а именно
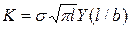 ,
, 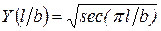 . (3.2.3.)
. (3.2.3.)
Здесь 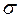 - приложенное к полосе растягивающее напряжение; l – полудлина трещины. Поскольку ширина полосы более чем в десять раз превышает длину трещины, то поправочный коэффициент
- приложенное к полосе растягивающее напряжение; l – полудлина трещины. Поскольку ширина полосы более чем в десять раз превышает длину трещины, то поправочный коэффициент  . При этом формула (3.2.3.) переходит в формулу для растянутой плоскости, которая имеет вид
. При этом формула (3.2.3.) переходит в формулу для растянутой плоскости, которая имеет вид
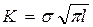 . (3.2.4)
. (3.2.4)
Подставляя это выражение в критериальное условие Ирвина
 (3.2.5)
(3.2.5)
получаем уравнение
 или
или  МПам1/2.
МПам1/2.
Отсюда искомое критическое окружное напряжение оказывается равным 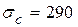 МПа, давление будет 3,3 МПа.
МПа, давление будет 3,3 МПа.
Видим, что разрушающее напряжение трубы с такой трещиной меньше предела текучести. Итак, на поставленный вопрос получен ответ.
Поинтересуемся теперь критической длиной трещины в случае, когда полоса растянута расчетным напряжением 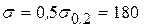 МПа (т.е.
МПа (т.е. 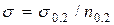 при коэффициенте запаса по пределу текучести
при коэффициенте запаса по пределу текучести  ). Условие разрушения (3.2.5) позволяет записать
). Условие разрушения (3.2.5) позволяет записать
 или
или 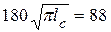 МПам1/2.
МПам1/2.
Отсюда критическая длина трещины, приводящая к полному разрушению полосы, равна 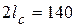 мм. Она оказалась больше, чем в первой части примера, поскольку действующее напряжение 180 МПа меньше, чем 290 МПа.
мм. Она оказалась больше, чем в первой части примера, поскольку действующее напряжение 180 МПа меньше, чем 290 МПа.
Таким образом, если теперь предположить, что стенка трубопровода растянута расчетным напряжением 180 МПа и в этой трубе есть трещина длиной 60 мм, то коэффициент запаса по разрушающему напряжению оказывается равным 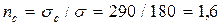 вместо двух по пределу текучести.
вместо двух по пределу текучести.
[1] Понятие трещиностойкости стоит в одном ряду с такими понятиями механики материалов,как пластичность, прочность, ползучесть и т.п. Эти понятия отражают явления, происходящие с материалом, и реакцию материала на внешнее воздействие. Мера количественной оценки этой реакции может быть измерена разными величинами. Например, для тела с трещиной характеристики трещиностойкости можно оценивать критическим коэффициентом интенсивности напряжений, пределом трещиностойкости, критическим раскрытием вершины трещины, удельной работой разрушения, критическим значением "джей"-интеграла, процентом волокна в изломе, критической температурой хрупкости, ударной вязкостью образца с трещиной и др.




