
ФУНКЦИОНИРОВАНИЯ БИОЛОГИЧЕСКИХ МЕМБРАН. ЯВЛЕНИЕ ПЕРЕНОСА.
1. Основные функции и строение биологических мембран.
Структурной единицей живого организма является клетка, выполняющая все основные жизненные функции. С помощью современных методов исследования структуры биологических объектов установлено, что важное значение в создании структуры клетки и ее функционирования имеют биологические мембраны. Мембраны, являясь универсальными структурами, выполняют многообразные и чрезвычайно важные для клетки функции: они окружают всю цитоплазму и ограничивают ее от окружающей среды, обеспечивают прочность и автономность клеток (механическая функция). Помимо мембраны, образующей наружный пограничный слой клетки, подобные мембранные структуры пронизывают цитоплазму и образуют оболочки всех клеточных органоидов: ядра, митохондрий, лизосом, аппарата Гольджи. Различного рода мембранные структуры в организмах животных и человека составляют колоссальную поверхность – десятки тысяч квадратных метров. Такая обширная структурная система указывает на ее важное функциональное значение. Мембраны регулируют обмен веществ клетки и служат ее осмотическим барьером (цитоплазматические мембраны), являются регулятором клеточного деления, играют большую роль в генерации и проведении потенциалов, в клеточном дыхании, являются местом локализации (служат основой, матрицей) для мембранных ферментов, макроэнергетических соединений, рецепторов и других встроенных в мембраны молекул, а также чувствительными приемниками и преобразователями световых, звуковых, механических и химических сигналов внешнего мира. Ряд жизненных процессов протекают на биологических мембранах. Многие болезни связаны с нарушением нормального функционирования мембран: канцерогенез, атеросклероз, отравление, вирусные и инфекционные заболевания, поражение организма УФ- и радиоактивным излучением. Поэтому лечение часто связано с воздействием на мембраны с целью нормализации их функций. Несмотря на то, что мембраны имеют различный химический состав и осуществляют специфические функции в различных клетках, в общих чертах они обладают универсальным строением. Теория молекулярного строения клеточных мембран вначале базировалась на некоторых косвенных данных.
В 1956 году модель была усовершенствована – введены в рассмотрение «поры» с тем, чтобы объяснить достаточно свободное проникновение через мембрану путем диффузии воды и таких гидрофильных соединений, как мочевина. Диаметр пор определяется косвенным путем по размерам водорастворимых молекул, которые еще способны проникать через мембрану внутрь клетки. С помощью этого и других методов было установлено, что диаметр пор составляет 0,35-0,8 нм. Поры могут иметь структуру длинного извитого канала. Количество пор в мембране не велико. В эритроцитах, например, вся площадь, приходящаяся на их долю, составляет примерно 0,06% от общей поверхности мембраны. Поры изнутри выстланы слоем молекул белка. Полярные группы молекул белка направлены в сторону отверстия поры, а неполярные вступают во взаимодействие с молекулами липидов. Благодаря наличию полярных групп в порах, они обычно обладают электрическим зарядом, что оказывает большое влияние на процесс проникновения растворимых частиц через поры, в частности, ионов. В дальнейшем было высказано множество гипотез о строении биологических мембран, но ни одна не стала общепринятой. В 1972 году Сингер и Николсон на основании результатов, полученных физическими и химическими методами исследования предложили модель, представляющую в настоящее время наиболее удовлетворительной - жидкостно-мозаичную, согласно которой фосфолипиды образуют двойной слой, но не обязательно непрерывный (двойной слой фосфолипидов, инкрустированный белками). Не вся поверхность биологической мембраны покрыта белками. Различают поверхностные (или периферические) и интегральные белки. За счет этих белков осуществляются важные функции мембраны: проницаемость, активный перенос через мембрану, генерация электрического потенциала. Липиды находятся в жидкой фазе. Белки включаются в слой липидов, но их полярные группы сохраняют контакт с водной фазой; есть белки, которые пронизывают мембрану насквозь, они формируют мембранные каналы, некоторые белки погружены в нее наполовину. Одни белки связаны друг с другом, другие в большей или меньшей степени окружены липидами (формально такое состояние можно сопоставить с гидротированными ионами). Распределение белков неравномерно. Согласно электронно-микроскопическим данным, концентрация их на внутренней поверхности выше, чем на наружной. Белки довольно подвижны, т.е. мембрана не является неподвижной структурой. Липиды и белки обмениваются местами, перемещаясь как вдоль ее плоскости, так и поперек - так называемый "флип-флоп". В состав мембран входят и другие химические соединения: холестерин, гликолипиды, гликопротеиды.
2. Липидные модельные мембраны. Природная мембрана очень сложная система, поэтому пытаются создавать различные модели, воспроизводящие ее структуру и свойства. Об организации молекул липидов в мембранах дают представления модели в виде искусственных мономолекулярных пленок (монослойная липидная мембрана - МЛМ). Техника получения искусственных плоских мембран была разработана Ленгмюром еще в 1917 году.
Вторая модель мембран - плоские бислойные липидные мембраны (БЛМ). Впервые такую мембрану создал П. Мюллер со своими сотрудниками в 1962 году. Для этого на отверстие в тефлоновой пластинке наносили раствор фосфолипидов в гептане. Затем пластинку помещали в раствор хлорида калия. По мере диффузии растворителя из капли в водную фазу отверстие оказывалось закрытым бислойной фосфолипидной мембраной толщиной 5-7 нм и диаметром немного более 1 мм. Такая мембрана может длительно существовать в водном растворе солей. Модели мембран такого рода сыграли большую роль в выяснении механизма действия веществ - ионофоров (например, антибиотиков типа валиномицин), а также ряда других соединений, модифицирующих проницаемость мембраны, в том числе, лекарственных препаратов, токсинов и т.д. На подобных моделях исследователи изучали электропроводность, транспорт через них ионов, их проницаемость для различных веществ, а также механические и оптические свойства.
Липосомы нашли применение в медицине. Например, можно включить внутрь липосомы лекарственный препарат и использовать ее как фосфолипидную микрокапсулу для доставки лекарства в определенные орган и ткани. Липосомы не токсичны (при правильном подборе липидов) и полностью усваиваются организмом. Основная ценность метода искусственных мембран заключается в том, что он позволяет изучить сложные биофизические процессы в мембранах, поскольку исследования проводятся на сравнительно простых системах. Он позволяет на искусственных мембранах исследовать процессы самоорганизации биологических структур, которые пока еще не полностью изучены.
3. Некоторые физические свойства мембран и методы их исследования. Основу биологической мембраны составляет двойной слой молекул фосфолипидов. Липидный компонент мембран определяет их следующие свойства: механические, оптические, электрические (R, C), осмотические (непроницаемость для ионов и проницаемость для воды). В мембране содержатся разные фосфолипиды. Например, в мембране эритроцитов их около 20. С электрической точки зрения мембрана представляет собой диэлектрик с относительной диэлектрической проницаемостью от 2 до 6. Экспериментально показано, что между двумя сторонами мембраны существует разность потенциалов 50-80 мВ. Учитывая толщину мембраны, эта разность потенциалов приводит к возникновению очень высокой напряженности порядка 104-105 В·см -1. Липиды составляют 20-30% сухого веса мембраны, при этом считается, что на одну молекулу белка приходится приблизительно 75-90 молекул липидов. Белки, входящие в состав мембран, весьма разнообразны. Молекула фосфолипидов состоит из двух функционально различных частей. Неполярный гидрофобный "хвост" (остатки жирных кислот) - длинная прямая углеводородная цепь (СН2; СН3), к которой не могут присоединяться молекулы воды. Эта углеводородная цепь присоединяется к более сложной структуре ("головке"), содержащей атомы углерода, водорода, кислорода, фосфора, азота. Эта фосфорсодержащая часть молекулы является электрически полярной и притягивает к себе молекулы воды, которые также являются полярными. Таким образом, один конец фосфолипидной молекулы обладает гидрофильными, а второй - гидрофобными свойствами. В головке фосфолипида имеются разноименно заряженные группы, расположенные на некотором расстоянии друг от друга, т.е. представляющие собой диполь. Биологические мембраны, толщина которых примерно равна 7-8 нм, представляют собой как бы два монослоя на поверхности раздела: вода-липид. Силой, сжимающей монослой в мембране, оказывается поверхностное натяжение на границе вода-липидная фаза. Это поверхностное натяжение имеет величину 0,03-1 мН/м и приводит к сжатию бислоя. Вязкость липидного слоя мембран на два порядка выше вязкости воды и равна 30-100 мПа·с (сравнима с вязкостью подсолнечного масла). Многие болезни связаны с отклонением микровязкости липидной фазы от нормы. Например, канцерогенез связан со снижением ее, а при старении организма вязкость увеличивается. Изменение состояния липидной молекулы, связанное с изменением температуры, химической модификацией жирнокислотного "хвоста" или же с изменением заряда головки, сопровождается изменением площади, занимаемой молекулами. К такому же результату приводит – взаимодействие мембран с лекарственными препаратами, например, с анестетиками. В биологических мембранах липиды находятся преимущественно в жидкокристаллическом состоянии. При изменении поверхностного натяжения жидкости на границе мембрана - окружающий раствор, поверхностного заряда или температуры в липидной фазе мембран могут происходить локальные или генерализованные фазовые переходы из жидкокристаллического в гель-состояние, которое условно иногда называют твердокристаллическим. Переходы обусловлены сложными физическими свойствами фосфолипидов, которые принадлежат к соединениям с длинными углеводородными цепочками. Они способны образовывать несколько кристаллических форм. Жидкокристаллическое состояние бислоя имеет меньшую вязкость, меньшую упорядоченность молекул, большую ионную проводимость, большую растворимость веществ, чем твердое состояние. Толщина жидкокристаллического бислоя меньше, чем твердокристаллического. Конформация (структура) молекул в жидком и твердом состоянии различна, в чем можно убедиться при рентгеноструктурном анализе. В жидкой фазе молекулы фосфолипидов могут образовывать полости ("кинки"), в которые способны внедряться молекулы диффундирующего вещества и которые способны перемещаться. Для исследования некоторых физических свойств биологических мембран используется метод флюоресцентного анализа (с использованием флюоресцентных зондов и меток). В нормальном состоянии мембрана не флюоресцирует. Чтобы произвести исследования, необходимо вводить в мембрану молекулы или молекулярные группы, способные к флюоресценции (ДМХ-диметиламинохалкон; АНС-анилин-нафталин-сульфонат и др.). Флюоресцентный анализ дает возможность исследовать подвижность фосфолипидных молекул в мембране, оценить вязкость липидной фазы (микровязкость мембран), по смещению флюоресценции в более короткую область спектра при увеличении вязкости. Микровязкость можно оценить по степени поляризации P флюоресцентного излучения при освещении мембраны поляризованным светом: Излучение флюоресценции оказывается лишь частично поляризованным (мембрана освещается полностью поляризованным светом). Чем больше подвижность молекулы (флюоресцирующей), тем меньше вязкость, тем меньше степень поляризации. Наиболее полное представление об агрегатном состоянии липидных бислоев дают методы радиоспектроскопии – электронный парамагнитный резонанс (ЭПР) с использованием метода спиновых зондов и ядерный магнитный резонанс (ЯМР).
Применение ЭПР основано на том, что форма кривой поглощения зависит от свойств окружения свободного радикала и в первую очередь от микровязкости среды (рис.4). В жидкокристаллической фазе жирнокислотные цепи фосфолипидов обладают значительной подвижностью, чем в твердой фазе. Этим методом было отмечено снижение подвижности при увеличении содержания холестерина, при перекисном окислении липидов, действии ряда лекарственных веществ. Увеличение подвижности отмечено при тиреотоксикозах и ряде других патологий. Но при этом методе изучается химически измененная молекула фосфолипидов. Изучение подвижности различных участков немодифицированных молекул фосфолипидов позволяет осуществить метод ядерного магнитного резонанса (ЯМР). Ядерный магнитный резонанс – это явление резкого возрастания поглощения энергии электромагнитной волны системой атомных парамагнитных ядер, помещенных в постоянное магнитное поле, при резонансной частоте волн νрез. В биологическом объекте содержится много парамагнитных ядер
4. Общее уравнение переноса. Диффузия. Уравнение Фика. Важным элементом функционирования мембран является их способность пропускать или не пропускать молекулы (атомы) и ионы.
Эти молекулы переносят через площадку свои физические характеристики. Рассмотрим общий механизм переноса, не конкретизируя, какую именно величину переносят молекулы и обозначим ее через j. Тогда количество физической характеристики, перенесенное молекулами в одном направлении через площадку DS за Dt:
Очевидно, что такое количество будет перенесено и в обратном направлении. Предположим теперь, что рассматриваемый газ неоднороден по своим свойствам, т.е. у него различная концентрация в различных местах и сами молекулы имеют неодинаковое значение j. Тогда количество физической величины, содержащейся в единице объема, также будет различно в разных местах. Пусть количество (jn0) убывает в положительном направлении OX, будучи равным (n0
Величину
Знак минус обусловлен тем, что перенос физической величины происходит в направлении, противоположном градиенту ( Выражение (5) называют уравнением переноса. На его основании рассмотрим конкретное явление переноса - диффузию, имеющую важное значение для проницаемости веществ в живых системах. Пусть плотность газа убывает в направлении оси ОX:
где ΔМ - масса газа, переносимая путем диффузии за время Δt через площадку ΔS, перпендикулярную направлению убывания плотности. Тогда: Уравнение Фика может быть записано через плотность потока вещества: ЛЕКЦИЯ № 18
|
 На основании анализа многочисленных исследований состава и свойств мембран Даниэлии и Давсон в 1935 году предложили модель строения биологической мембраны, которая, в основном, не претерпела существенных изменений до нашего времени. Согласно этой модели мембрана имеет структуру сэндвича – двойной липидный слой зажат между слоями белков (рис.1). Липидные молекулы в слоях расположены перпендикулярно поверхности мембраны, гидрофильные концы молекул липидов направлены наружу, а гидрофобные – к центру мембраны. Наличие липидов в структуре биологических мембран подкрепляется результатами полученными при измерении электрических параметров клетки, которые свидетельствуют о высоком сопротивлении клеточной мембраны (порядка 1000 Ом/см2), характерной для липидов, и значительной емкостью (0,05 мкФ/м2). Липиды связаны друг с другом гидрофобными взаимодействиями, а липиды и белки электростатическими силами. Белковые молекулы в форме глубул покрывают двойной слой фосфолипидов с обоих сторон, придавая ему тем самым определенную эластичность, устойчивость к механическим повреждениям, а так же низкое поверхностное натяжение (0,1 дин/см). Полярные группы молекул белков направлены в сторону водной фазы, а неполярные – в сторону липидов.
На основании анализа многочисленных исследований состава и свойств мембран Даниэлии и Давсон в 1935 году предложили модель строения биологической мембраны, которая, в основном, не претерпела существенных изменений до нашего времени. Согласно этой модели мембрана имеет структуру сэндвича – двойной липидный слой зажат между слоями белков (рис.1). Липидные молекулы в слоях расположены перпендикулярно поверхности мембраны, гидрофильные концы молекул липидов направлены наружу, а гидрофобные – к центру мембраны. Наличие липидов в структуре биологических мембран подкрепляется результатами полученными при измерении электрических параметров клетки, которые свидетельствуют о высоком сопротивлении клеточной мембраны (порядка 1000 Ом/см2), характерной для липидов, и значительной емкостью (0,05 мкФ/м2). Липиды связаны друг с другом гидрофобными взаимодействиями, а липиды и белки электростатическими силами. Белковые молекулы в форме глубул покрывают двойной слой фосфолипидов с обоих сторон, придавая ему тем самым определенную эластичность, устойчивость к механическим повреждениям, а так же низкое поверхностное натяжение (0,1 дин/см). Полярные группы молекул белков направлены в сторону водной фазы, а неполярные – в сторону липидов. Если на поверхность воды нанести каплю растворенных в каком-либо летучем растворителе фосфолипидов, то после распределения их молекул по водной поверхности и испарения растворителя образуется мономолекулярная пленка (рис.2). При полном насыщении поверхностного слоя адсорбированные молекулы липидов располагаются перпендикулярно к поверхности воды таким образом, что в воду погружается гидрофильная (полярная) головка, а неполярная углеводородная цепь направлена вертикально вверх. Такой ориентированный слой молекул называют "частоколом Ленгмюра". Толщина такой мономолекулярной пленки в случае стеариновой кислоты составляет 2,5 нм. Молекулы фосфолипидов в таком монослое располагаются так же плотно, как в одном из бислоев мембраны. Если стеклянную пластинку опустить в воду, на поверхности которой находится мономолекулярная пленка, то эту пленку можно перенести на поверхность пластинки. При повторных погружениях на пластинке возникают бимолекулярные пленки.
Если на поверхность воды нанести каплю растворенных в каком-либо летучем растворителе фосфолипидов, то после распределения их молекул по водной поверхности и испарения растворителя образуется мономолекулярная пленка (рис.2). При полном насыщении поверхностного слоя адсорбированные молекулы липидов располагаются перпендикулярно к поверхности воды таким образом, что в воду погружается гидрофильная (полярная) головка, а неполярная углеводородная цепь направлена вертикально вверх. Такой ориентированный слой молекул называют "частоколом Ленгмюра". Толщина такой мономолекулярной пленки в случае стеариновой кислоты составляет 2,5 нм. Молекулы фосфолипидов в таком монослое располагаются так же плотно, как в одном из бислоев мембраны. Если стеклянную пластинку опустить в воду, на поверхности которой находится мономолекулярная пленка, то эту пленку можно перенести на поверхность пластинки. При повторных погружениях на пластинке возникают бимолекулярные пленки. Третьей модельной системой, которая широко используется в настоящее время, является липосомы. Липосомы - это фосфолипидные пузырьки (везикулы). Они могут быть получены путем встряхивания сухих фосфолипидов в водно-солевом буфере. Суспензия липосом - очень удобный объект для изучения структуры липидов бислоя (рис.3), т.к. строение этого слоя близко к строению в биологических мембранах, мембранах митахондрий, эндоплазматического ретикулума, эритроцитов и др. клеток.
Третьей модельной системой, которая широко используется в настоящее время, является липосомы. Липосомы - это фосфолипидные пузырьки (везикулы). Они могут быть получены путем встряхивания сухих фосфолипидов в водно-солевом буфере. Суспензия липосом - очень удобный объект для изучения структуры липидов бислоя (рис.3), т.к. строение этого слоя близко к строению в биологических мембранах, мембранах митахондрий, эндоплазматического ретикулума, эритроцитов и др. клеток. , где Jпол. – интенсивность поляризованного света; Jсумм. – суммарная интенсивность флюоресценции.
, где Jпол. – интенсивность поляризованного света; Jсумм. – суммарная интенсивность флюоресценции. ЭПР – это явление резкого возрастания поглощения энергии электромагнитного излучения (микроволновый диапазон) системой парамагнитных частиц (обладают не спаренными электронами и не скомпенсированным магнитным моментом), помещенных в постоянное магнитное поле, при резонансной частоте волны νрез.. Спектром ЭПР называют зависимость энергии (E) поглощения электромагнитной волны от величины магнитной индукции B (рис. 4). При помощи ЭПР можно изучать лишь объекты, обладающие не спаренными электронами (свободные радикалы). Поэтому в изучаемую систему, производя соответствующий химический синтез, вводят спиновые метки - (спин-зонды) - соединения представляющие собой различные нитроксильные радикалы (
ЭПР – это явление резкого возрастания поглощения энергии электромагнитного излучения (микроволновый диапазон) системой парамагнитных частиц (обладают не спаренными электронами и не скомпенсированным магнитным моментом), помещенных в постоянное магнитное поле, при резонансной частоте волны νрез.. Спектром ЭПР называют зависимость энергии (E) поглощения электромагнитной волны от величины магнитной индукции B (рис. 4). При помощи ЭПР можно изучать лишь объекты, обладающие не спаренными электронами (свободные радикалы). Поэтому в изучаемую систему, производя соответствующий химический синтез, вводят спиновые метки - (спин-зонды) - соединения представляющие собой различные нитроксильные радикалы ( ), которые можно присоединить к любому атому углерода углеводородной цепочки молекулы липида.
), которые можно присоединить к любому атому углерода углеводородной цепочки молекулы липида. - протонов, что дает возможность применять их для исследования методом ЯМР. При ЯМР частота переменного электромагнитного поля меньше, чем при ЭПР.
- протонов, что дает возможность применять их для исследования методом ЯМР. При ЯМР частота переменного электромагнитного поля меньше, чем при ЭПР. Проницаемость обусловлена целым рядом важных физических явлений и, в частности, переносом молекулами веществ (в процессе движения) своих физических характеристик: массы, энергии, импульса и т.д. Механизм переноса всех этих характеристик одинаков и все явления, связанные с их переносом, объединены общим названием явления переноса. Исходя из представлений молекулярно-кинетической теории, выведем общее для явлений переноса уравнение переноса (на примере газа). С этой целью определим прежде всего количество молекул, переходящих за промежуток времени ∆t через некоторую воображаемую площадку ∆S, помещенную в газе. Ось ОX ориентирована перпендикулярно ∆S (рис.5). Через площадку проходит 1/3 всех молекул, движущихся вдоль ОX (1/6 – слева направо; 1/6 - справа налево). Слева направо за 1 секунду пройдет через DS 1/6 всех молекул, находящихся в объеме прямоугольного параллелепипеда с основанием DS и высотой, равной средней скорости движения молекул
Проницаемость обусловлена целым рядом важных физических явлений и, в частности, переносом молекулами веществ (в процессе движения) своих физических характеристик: массы, энергии, импульса и т.д. Механизм переноса всех этих характеристик одинаков и все явления, связанные с их переносом, объединены общим названием явления переноса. Исходя из представлений молекулярно-кинетической теории, выведем общее для явлений переноса уравнение переноса (на примере газа). С этой целью определим прежде всего количество молекул, переходящих за промежуток времени ∆t через некоторую воображаемую площадку ∆S, помещенную в газе. Ось ОX ориентирована перпендикулярно ∆S (рис.5). Через площадку проходит 1/3 всех молекул, движущихся вдоль ОX (1/6 – слева направо; 1/6 - справа налево). Слева направо за 1 секунду пройдет через DS 1/6 всех молекул, находящихся в объеме прямоугольного параллелепипеда с основанием DS и высотой, равной средней скорости движения молекул  , т.е. 1/6ΔS
, т.е. 1/6ΔS 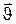 n0, где n0 - число молекул в единице объема. За время Dt пройдет:
n0, где n0 - число молекул в единице объема. За время Dt пройдет: . (1)
. (1) . (2)
. (2) )1 слева от площадки ΔS и (n0
)1 слева от площадки ΔS и (n0 
 . (3)
. (3) Теперь остаётся только выяснить, на каком расстоянии от ΔS следует взять значение
Теперь остаётся только выяснить, на каком расстоянии от ΔS следует взять значение  . Обмен значениями
. Обмен значениями  (средняя длина свободного пробега). Можно полагать, что
(средняя длина свободного пробега). Можно полагать, что  слева и справа от площадки. На этих расстояниях будем брать значение
слева и справа от площадки. На этих расстояниях будем брать значение  . Умножив и разделив на
. Умножив и разделив на  правую часть формулы (3), получим:
правую часть формулы (3), получим: . (4)
. (4) - называют градиентом
- называют градиентом  . (5)
. (5) направлен справа налево, а перенос
направлен справа налево, а перенос  - плотности на расстоянии
- плотности на расстоянии  (рис. 7). Так как
(рис. 7). Так как  , где m - масса молекулы, то n01>n02.
, где m - масса молекулы, то n01>n02. Учитывая в уравнении (5), что переносимой величиной будет масса молекулы, т.е.
Учитывая в уравнении (5), что переносимой величиной будет масса молекулы, т.е. 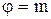 , получим:
, получим:  ;
;  ,
, . Обозначим
. Обозначим  и запишем:
и запишем:  , где D - коэффициент диффузии, размерность которого [м2/с]. Откуда следует, что масса газа ΔМ, переносимая благодаря диффузии через площадку ΔS, перпендикулярную оси Ох, в направлении которой убывает плотность, пропорциональна площади этой площадки, промежутку времени Δt и градиенту плотности Δρ/Δx (уравнение диффузии, закон Фика).
, где D - коэффициент диффузии, размерность которого [м2/с]. Откуда следует, что масса газа ΔМ, переносимая благодаря диффузии через площадку ΔS, перпендикулярную оси Ох, в направлении которой убывает плотность, пропорциональна площади этой площадки, промежутку времени Δt и градиенту плотности Δρ/Δx (уравнение диффузии, закон Фика). ,
, 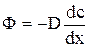 ; где
; где  - градиент концентрации.
- градиент концентрации.


