
БИОЭЛЕКТРИЧЕСКИЕ ПОТЕНЦИАЛЫ.
1. Мембранные потенциалы и их ионная природа. Все процессы жизнедеятельности организма сопровождаются появлением в клетках и тканях электрических потенциалов, которые получили название биоэлектрических. Это явление лежит в основе возбудимости клетки, регуляции внутриклеточных процессов, работы нервной системы, регуляции мышечного сокращения. При этом имеется в виду разность потенциалов между двумя точками живой системы. Такие разности имеют место между содержимым живой клетки и окружающей её средой, между возбуждённым и невозбуждённым участками одной и той же клетки, между участками одного и того же органа, находящимися в различных физиологических состояниях. Благодаря непосредственной связи биопотенциалов с метаболическими процессами и физиологическим состоянием клетки, они являются весьма чувствительными и точно измеряемыми показателями различных изменений в клетках в норме и патологии. Нарушение электрических процессов в клетках и тканях приводит к ряду серьёзных патологий или сопутствует заболеваниям. Исследование электрических потенциалов головного мозга, сердца, мышц, глаза и других органов даёт возможность установить не только расстройства в деятельности этих органов, но и в значительной мере выявить причину этих расстройств. Эти исследования лежат в основе диагностических методов: энцефалографии, электрокардиографии, электромиографии, электроретинографии. Причина наличия разности потенциалов в живых системах кроется в существовании определённых физико-химических градиентов между отдельными тканями, между клеточным содержимым и окружающей клетку жидкостью. Различают окислительно-восстановительные потенциалы, возникающие вследствие переноса электронов от одной молекулы к другой. Существуют и мембранные потенциалы, которые возникают вследствие градиента концентрации ионов и переноса их через мембрану, причём один из растворов содержит ионы, или плохо пропускаемые мембраной, или совсем ей не пропускаемые. Биопотенциалы, регистрируемые в организме, – это, в основном, мембранные. Они доступны для измерения и используются в диагностических целях. Мембранный потенциал обусловлен диффузией, однако, диффузией не в свободном пространстве, а через мембрану, когда различия в подвижности анионов и катионов могут значительно увеличиваться. Мембранным потенциалом называется разность потенциалов между внутренней (цитоплазматической) и наружной поверхностью мембраны:
где C1 – концентрация ионов в области, откуда идёт диффузия; С2 – концентрация ионов в области, куда идёт диффузия; T – абсолютная температура; F – число Фарадея (96500 Кл/моль); Z – заряд иона (в единицах элементарного заряда); R – универсальная газовая постоянная (8,3 Дж/Кл · моль). Как следует из уравнения, мембранный потенциал зависит от температуры и от величины концентрационного градиента диффундирующих через мембрану ионов (остальные величины постоянные). Нечто сходное с указанным примером имеет место в живой клетке, в которой концентрация ионов калия больше, чем в окружающей межклеточной жидкости. Мембранная теория биопотенциалов хорошо теоретически разработана и подтверждена блестящими экспериментами. Она была выдвинута ещё в 1902 г. Бернштейном. Однако только в 50-х годах эта теория была по настоящему развита и экспериментально обоснована Ходжкиным, которому принадлежат основные идеи и теории о роли ионных градиентов в возникновении потенциалов и о механизме распределения ионов между клеткой и средой. 2. Потенциал покоя. Потенциал покоя – это стационарная разность электрических потенциалов, регистрируемых между наружной и внутренней поверхностями мембраны в невозбужденном состоянии. Потенциал покоя определяется разной концентрацией ионов по разным сторонам мембраны и диффузией ионов через мембрану.
Согласно современным взглядам, причиной возникновения потенциала клеток в покое является неравномерное распределение ионов калия между содержимым клетки и окружающей средой. Концентрация ионов калия в клетках в 20-40 раз превышает их содержимое в окружающей среде. Эта точка зрения базируется на том, что ионы калия внутри клетки находятся в свободном состоянии и могут легко диффундировать. Исследования Ходжкина и Кейнеса (1953г.), выполненные с радиоактивным 42К на гигантских аксонах кальмара, показали, что подвижность и коэффициент диффузии ионов 42К внутри волокна почти такие же, как в водном растворе. Избыток положительных зарядов ионов калия внутри клеток компенсируется в основном органическими анионами, содержание которых в клетках сравнительно небольшое. К ним относятся анионы аспарагиновой, уксусной, пировиноградной и других органических кислот. Согласно Ходжкина, Хаксли, Катца клеточная мембрана проницаема в состоянии покоя только для ионов калия. Ионы калия диффундируют по градиенту концентрации через клеточную мембрану в окружающую жидкость, анионы не могут проникать через клеточную мембрану и остаются на ее внутренней стороне. Внешняя поверхность мембраны при этом заряжается положительно, а внутренняя отрицательно. Если принять, что потенциал покоя определяется диффузией только ионов калия из цитоплазмы наружу, то его величина Разность потенциалов между внутренней и внешней поверхностью мембраны различных клеток, рассчитанная по уравнению Нернста, оказалось очень близкой к измеренной в эксперименте с помощью внутриклеточных микроэлектродов. Кроме того, калиевый механизм возникновения потенциала покоя убедительно подтверждают эксперименты с изменением концентрации калия в окружающей клетку среде. При изменение наружной концентрации калия в широких пределах наблюдалось закономерное изменение величины потенциалов покоя в соответствии с зависимостью, выраженной уравнением Нернста. В частности, когда концентрацию калия в окружающей клетку среде делали равной концентрации его в клетке, потенциал покоя становился равным нулю. При измерении потенциала покоя выяснилось, что в одних случаях его измеренное значение соответствует значению, теоретически вычисленному по уравнению Нернста, в других случаях между измеренной и вычисленной величинами имеются значительные различия. Это объясняется тем, что на величину потенциала покоя, наряду с ионами К+, оказывают влияние другие ионы, проникающие через мембрану, и показывает, что «калиевая теория» потенциала покоя несовершенна и не полностью объясняет наблюдаемые факты. Для количественного описания в условиях проницаемости мембран для нескольких ионов Ходжкин и Катц использовали представление о том, что потенциал покоя не равновесный, а стационарный по своей природе, т.е. он отражает состояние системы, когда через мембрану непрерывно идут встречные потоки К+, Na+,Cl-. Суммарная плотность потока равна сумме плотности положительно заряженных ионов (одновалентных катионов) минус сумма плотности потоков одновалентных анионов: Ф=ФК++ФNa+-ФCl-. Перед ФCl – стоит знак минус, учитывающий отрицательный заряд иона хлора. В стационарном состоянии суммарная плотность потока равна нулю (Ф=0), т.е. число различных ионов, проходящих в единицу времени через мембрану внутрь клетки, равно числу выходящих из клетки через мембрану ионов. Выразив плотности потоков положительных и отрицательных ионов, просуммируем их и приравниваем к нулю, получим:
Третий член этого выражения соответствует плотности потока отрицательно заряженных ионов Cl-. После ряда преобразований можно получить выражение для потенциала покоя на мембране для 3-х ионов:
Это уравнение называется уравнением стационарного потенциала Гольдмана-Ходжкина-Катца или проще – уравнением Гольдмана. Последнее переходит в уравнение Нернста, если проницаемость одного из ионов гораздо выше, чем для других. Например, в гигантской аксоне кальмара РК:РNa:РCl=1:0,04:0,45, т.е. РК заметно выше, чем для других ионов, поэтому числитель и знаменатель в уравнении примерно равны соответственно
3. Потенциал действия.
Потенциал действия – это электрический импульс, обусловленный изменением ионной проводимости мембраны в связи с распространением по нервам и мышцам волны возбуждения. Опыты по исследованию потенциала действия проведены (в основном Хаджкиным и его сотрудниками) на гигантских аксонах кальмара методом микроэлектродов, а так же методом меченых атомов. Клетки различных тканей при действии различных раздражителей (механических, тепловых, электрических) способны переходить в состояние возбуждения. Возбуждаемость – это способность клеток к быстрому ответу на раздражение, проявляющемуся в совокупности физических, физико-химических процессов и функциональных изменений. Обязательным признаком возбуждения является изменение электрического состояния клеточной мембраны. Опыт показывает, что возбужденный участок клетки становится электроотрицательным по отношению к невозбужденному, что показывает на перераспределение ионов в возбужденном участке. Это перераспределение ионов носит временный характер и после окончания возбуждения вновь восстанавливается потенциал покоя. Характерной особенностью возникновения потенциала действия является необходимость сверхпорогового возбуждения. Возникновение возбуждения в живых системах сопровождается изменениями проникающей способности их клеточных мембран. В момент возбуждения резко меняется сопротивление мембраны, например, у аксона кальмара от 0,1 0м∙м2 в покое до 0,0025 Ом·м2 при возбуждении. Было показано, что возникновение потенциала действия связано с увеличением проницаемости мембран для ионов натрия и последующим усилением диффузии этих ионов по концентрационному градиенту внутрь клетки, что приводит к изменению (уменьшению) мембранного потенциала или деполяризации мембраны. Уменьшение мембранного потенциала ниже критического уровня приводит к еще большему увеличению проницаемости мембраны для натрия, а увеличение проницаемости сопровождается усилением диффузии Na+ в цитоплазму, что вызывает еще большую деполяризацию мембраны. По данным Ходжкина отношение коэффициента проницаемости мембраны аксона кальмара для фазы деполяризации мембраны: PK:PNa:PCl=1:20:0,45. Если сравнивать его с аналогичным соотношением в состоянии ионов:PK:PNa:PCl=1:0,04:0,45, то видно, что для К+ и Сl- в первой фазе возбуждения проницаемость не изменяется, а для Na+ она увеличивается в 500 раз. Усиленный поток положительно заряженных ионов Na+ внутрь клетки вызывает вначале исчезновение избыточного отрицательного заряда на внутренней поверхности мембраны (фаза деполяризации), а затем приводит к перезарядке мембраны, т.е. наружная поверхность мембраны на участке возбуждения становится отрицательной по отношению к невозбужденному. Эта фаза развития потенциала действия называется фазой реверсии. Поступление ионов натрия в клетку продолжается до тех пор, пока внутренняя поверхность мембраны не приобретет положительный заряд, достаточный для уравновешивания градиента концентрации Na+ и прекращения его перехода внутрь клетки. Повышенная проницаемость мембраны для ионов натрия продолжается очень короткий интервал времени (0,5-1мс). Затем усиливается поток ионов К+ из клетки, что приводит к восстановлению потенциала покоя на мембране, т.е. ее наружная поверхность за счет вышедших наружу ионов К+ опять приобретает положительный заряд, а внутренняя – отрицательный. Эта фаза, в течение которой поляризация мембраны возвращается на исходный уровень, называется фазой реполяризации. Фаза реполяризации всегда продолжительнее фазы деполяризации. Таким образом, восстановление потенциала покоя происходит не в результате обратного перемещения ионов Na+, а вследствие выхода эквивалентного количества ионов К+. В период покоя нормальное состояние концентраций восстанавливается, что обуславливается работой калий-натриевого насоса, который обеспечивает активный перенос этих ионов.
На рис.3: АВ – фаза деполяризации; ВС – фаза реверсии; СД – фаза реполяризации. Таким образом, на основе обобщения большого экспериментального материала было установлено, что потенциал действия возникает в результате избыточной по сравнению с покоем диффузией ионов натрия из окружающей среды в клетку. Фактически, формирование потенциала действия обусловлено двумя ионными потоками через мембрану. Потоки ионов Na+ и К+ примерно равны по величине, но сдвинуты по времени. Благодаря этому сдвигу во времени и возможно возникновение потенциала действия. Если бы потоки натрия и калия через мембрану совпадали во времени, то они бы компенсировали друг друга, и никакого изменения мембранного потенциала не могло бы происходить. Характерной особенностью потенциала действия является период рефрактерности (невозбуждения мембраны) во время его развития и остаточные явления в течение 1-3 мс после снятия возбуждения. Это свидетельствует о том, что проницаемость мембраны окончательно восстанавливается не сразу после окончания потенциала действия. 4. Распространение потенциала действия по нервному волокну.
Локальный ток вызывает увеличение проницаемости соседних невозбужденных участков мембраны, что снижает их потенциал покоя по абсолютной величине, т.е. потенциал повышается. Когда деполяризация достигнет критического (порогового) значения, в этом участке возникает потенциал действия. На участке ранее возбужденном в это время происходят восстановительные процессы реполяризации. Этот процесс многократно повторяется и обеспечивает распространение импульса возбуждения дальше на покоящиеся участки. Под влиянием локальных токов волна возбуждения распространяется без затухания в одном направлении. Может возникнуть вопрос, почему возбуждение распространяется по аксону не в обе стороны от зоны, до которой дошло возбуждение (ведь локальные токи текут в обе стороны от возбужденного участка). Дело в том, что возбуждение может распространяться в области мембраны, находящейся в состоянии покоя, т.е. в одну сторону от возбужденного участка аксона. В другую сторону нервный импульс не может распространяться, так как области, через которые прошло возбуждение, некоторое время остаются невозбудимыми - рефрактерными. Нервные импульсы проводятся без затухания по аксонам в какой-то степени аналогично передаче электрических сигналов по кабельно-релейной линии.
ЛЕКЦИЯ №20
|
 . (1)
. (1) Рассмотрим пример возникновения потенциала на мембране (полупроницаемой). Если сосуд, изображённый на рис.1, разделить мембраной, не пропускающей органические соединения, и оба отсека заполнить раствором KCl, а затем в отсек 2 добавить калиевую соль гемоглобина (KHb), то ионы калия будут переходить из правой половины сосуда в левую, вследствие разности их концентрации (C1>C2). Отрицательные ионы
Рассмотрим пример возникновения потенциала на мембране (полупроницаемой). Если сосуд, изображённый на рис.1, разделить мембраной, не пропускающей органические соединения, и оба отсека заполнить раствором KCl, а затем в отсек 2 добавить калиевую соль гемоглобина (KHb), то ионы калия будут переходить из правой половины сосуда в левую, вследствие разности их концентрации (C1>C2). Отрицательные ионы  концентрируются на правой стороне мембраны и удерживают положительные ионы калия слева. Мембрана поляризуется, на ней возникает мембранный потенциал, определяемый на основании следующего уравнения:
концентрируются на правой стороне мембраны и удерживают положительные ионы калия слева. Мембрана поляризуется, на ней возникает мембранный потенциал, определяемый на основании следующего уравнения: 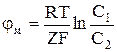 , (2)
, (2) В покоящейся, нормально функционирующей клетке всегда имеется разность потенциалов между цитоплазмой и окружающей средой. Еще в 1838 г. Маттеучи впервые установил, что наружная поверхность мышц заряжена положительно, а внутренняя – отрицательно. Эта разность потенциалов, свойственная состоянию покоя, и получила название потенциала покоя или мембранного потенциала. Определенное время считали, что потенциал покоя возникает вследствие повреждения клетки или ткани между поврежденным и неповрежденным участками. Поврежденный участок приобретал стойкий отрицательный заряд по отношению к неповрежденному. Однако было обнаружено, что разность потенциалов может возникать и у неповрежденных клеток. Экспериментально величина потенциала покоя была измерена с помощью микроэлектродов, один из которых вводился внутрь клетки (рис.2), а второй помещался во внеклеточную жидкость (1). Если оба электрода поместить во внеклеточную жидкость, то прибор не зафиксирует разность потенциалов (2).
В покоящейся, нормально функционирующей клетке всегда имеется разность потенциалов между цитоплазмой и окружающей средой. Еще в 1838 г. Маттеучи впервые установил, что наружная поверхность мышц заряжена положительно, а внутренняя – отрицательно. Эта разность потенциалов, свойственная состоянию покоя, и получила название потенциала покоя или мембранного потенциала. Определенное время считали, что потенциал покоя возникает вследствие повреждения клетки или ткани между поврежденным и неповрежденным участками. Поврежденный участок приобретал стойкий отрицательный заряд по отношению к неповрежденному. Однако было обнаружено, что разность потенциалов может возникать и у неповрежденных клеток. Экспериментально величина потенциала покоя была измерена с помощью микроэлектродов, один из которых вводился внутрь клетки (рис.2), а второй помещался во внеклеточную жидкость (1). Если оба электрода поместить во внеклеточную жидкость, то прибор не зафиксирует разность потенциалов (2).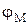 может быть найдена из уравнения Нернста для равновесного мембранного потенциала:
может быть найдена из уравнения Нернста для равновесного мембранного потенциала: 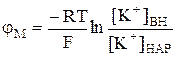 . (3)
. (3)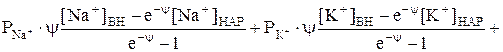
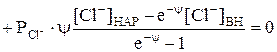
 .
.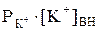 и
и  , благодаря чему уравнение превращается в уравнение Нернста для К+, т.е. уравнение Нернста является частным случаем уравнения Гольдмана. Рассчитанный по уравнению Гольдмана потенциал гигантского аксона кальмара совпадает с найденным в эксперименте.
, благодаря чему уравнение превращается в уравнение Нернста для К+, т.е. уравнение Нернста является частным случаем уравнения Гольдмана. Рассчитанный по уравнению Гольдмана потенциал гигантского аксона кальмара совпадает с найденным в эксперименте. Потенциалы действия представляют собой кратковременные быстро изменяющиеся потенциалы. Длительность потенциала действия в нервных волокнах составляет 1 мс, в сердечной мышце около 300 мс. Величина потенциала действия нерва равна 100-110 мВ, для скелетной мышцы – 120 мВ. Изменение потенциала на мембране можно представить графически на примере гигантского аксона кальмара (рис.3). Так в аксоне кальмара потенциал покоя равен – 45мВ и внутренняя поверхность мембраны электроотрицательная. При возбуждении потенциал внутренней поверхности мембраны становится равным +40мВ и происходит инверсия знака потенциала. Следовательно, полное значение потенциала действия составляет 85мВ, что значительно превышает потенциал покоя.
Потенциалы действия представляют собой кратковременные быстро изменяющиеся потенциалы. Длительность потенциала действия в нервных волокнах составляет 1 мс, в сердечной мышце около 300 мс. Величина потенциала действия нерва равна 100-110 мВ, для скелетной мышцы – 120 мВ. Изменение потенциала на мембране можно представить графически на примере гигантского аксона кальмара (рис.3). Так в аксоне кальмара потенциал покоя равен – 45мВ и внутренняя поверхность мембраны электроотрицательная. При возбуждении потенциал внутренней поверхности мембраны становится равным +40мВ и происходит инверсия знака потенциала. Следовательно, полное значение потенциала действия составляет 85мВ, что значительно превышает потенциал покоя. Потенциал действия, возникнув в одном возбужденном участке нервного волокна, быстро распространяется по всей его поверхности. Распространение потенциала действия обусловлено возникновением так называемых локальных токов, циркулирующих между возбужденным и невозбужденным участками. Рассмотрим распространение возбуждения на примере передачи нервного импульса по аксону. В состоянии покоя внешняя поверхность мембраны имеет положительный заряд, а внутренняя – отрицательный. В момент возбуждения полярность мембраны (на возбужденном участке) меняется на отрицательную (рис.4). В результате этого между возбужденным и невозбужденным участками возникает разность потенциалов, что приводит к появлению электрических токов между этими участками (локальные токи или токи действия). На поверхности мембраны ток течет от невозбужденного участка к возбужденному, а внутри – в противоположном направлении.
Потенциал действия, возникнув в одном возбужденном участке нервного волокна, быстро распространяется по всей его поверхности. Распространение потенциала действия обусловлено возникновением так называемых локальных токов, циркулирующих между возбужденным и невозбужденным участками. Рассмотрим распространение возбуждения на примере передачи нервного импульса по аксону. В состоянии покоя внешняя поверхность мембраны имеет положительный заряд, а внутренняя – отрицательный. В момент возбуждения полярность мембраны (на возбужденном участке) меняется на отрицательную (рис.4). В результате этого между возбужденным и невозбужденным участками возникает разность потенциалов, что приводит к появлению электрических токов между этими участками (локальные токи или токи действия). На поверхности мембраны ток течет от невозбужденного участка к возбужденному, а внутри – в противоположном направлении. ). Эти скорости распространения у некоторых беспозвоночных достигают 20-30 м/с и обеспечиваются большим диаметром (до 1 мм) их волокон. Нарушение миелиновой оболочки приводит к нарушению распространения потенциала действия по нервному волокну, и, как следствие, к тяжелым нервным заболеваниям.
). Эти скорости распространения у некоторых беспозвоночных достигают 20-30 м/с и обеспечиваются большим диаметром (до 1 мм) их волокон. Нарушение миелиновой оболочки приводит к нарушению распространения потенциала действия по нервному волокну, и, как следствие, к тяжелым нервным заболеваниям.


