
Регуляция остеогенеза, минерализации и деминерализации костной ткани.
Кость состоит из так называемых основных многоклеточных единиц ремоделирования, ответственных за локальные формообразование и местные концентрации кальция и фосфора. В составе таких единиц имеются мононуклеарные потомки недифференцированных мезенхимальных клеток – остеобласты. Они синтезируют коллаген 1 типа, располагают рецепторами паратгормона и ответственны за отложение органического остеоида и его последующую минерализацию. Маркером их активности служит секретируемый ими фермент – щелочная фосфатаза. Минерализация обеспечивается при участии минорных неколлагеновых кальций-связывающих белков остеобластов, которые содержат остатки α-карбоксиглютаминовой кислоты, фиксирующей кальций. К ним относятся остеокальцин и мофогенетический белок кости (матриксный кабоксиглютамил-содержащий белок, Gla-протеин матрикса). Карбоксиглютаминирование обоих белков зависит от витамина К.. Остеокальцин уникален для костей и зубов и его уровень в крови отражает скорость остеогенеза. Параллельно, через тромбоспондин, остеонектин и остеопонтин, эти фиксаторы кальция (и магния) закрепляются на коллагеновой матрице. Окружая себя минерализованным остеоидом, остеобласты превращаются в остеоциты, цитоплазма которых образует отростки, через гаверсовы канальцы остеоида связанные с соседними остеоцитами. Остеоциты участвуют в локальной перилакунарной деструкции кости и могут влиять на быстрые колебания уровня кальция в крови. Однако, основную остеолитическую
функцию в единицах ремоделирования кости выполняют потомки моноцитов – гигантские многоядерные макрофаги костей – остеокласты. Остеокласты перемещаются и образуют в участках резорбируемой кости, в особых лакунах Хоушипа, активный слой, прикрепляясь через специальный адаптер – αgβ3-интегрин – к остеопонтину. Они выделяют на своей активной гофрированной каёмке коллагеназу и маркерный фермент – кислую фосфатазу, лизируя минерализованный остеоид и растворяя кристаллы гидроксиапатита. Для этого, с помощью специальных протонного АТФазного насоса и карбоангидразы 2 типа, ими локально создаётся зона кислого рН=4. Молодой неминерализованный остеоид устойчив к их действию. Повреждённая кость при воспалении резорбируется ими и заменяется остеобластами на новую. Молодые остеокласты имеют рецепторы парат-гормона и кальцитонина, но на зрелых остаются лишь последние. Нет у них и рецепторов кальцитриола. Дифференцировка остеокластов зависит от гранулоцитарно-моноцитарного колониестимулирующего фактора, интерлейкина ИЛ-6 и паратгормона. Остеобласты и остеокласты функционируют согласованно (рис 2.9), что приводит к обновлению всего кальция костей за период, примерно, в 5-6 лет. Рост костей в длину зависит от энхондрального образования костной ткани на месте метаэпифизарного хряща, а в ширину (толщину) – от периостального окостенения. Костная ткань находится под контролем многих гормонов. Так, СТГ, пролактин, инсулин и андрогены способствуют синтезу остеоида. Инсулин стимулирует пролиферацию клеток и синтез в них КЛ1 и НКБ. Потенцирует действие фактор роста скелета (ФРС). (Установлено, что инсулин - необходимый компонент всех сред для культивирования клеток и тканей, т.е. роль в репликации и росте клеток несомненна). Глюкагон стимулирует секрецию кальцитонина (КТ). Глюкокортикоиды снижают в костях синтез коллагена, а также, препятствуя действию КТ в кишечнике и, уменьшая почечную реабсорбцию кальция, способствуют потере этого иона и остеопорозу. Эстрогены способствуют синтезу остеоида и отложению кальция в костях, как опосредованно через главные регуляторы кальциевого обмена, так и непосредственно. Андрогены (наиболее активный представитель - тестостерон) вызывают общий анаболический эффект. Индуцируют синтез белка в хондроцитах хрящевой зоны роста и в остеобластах, что сопровождается задержкой Са2+ и Р043- и увеличением массы костной ткани. Анаболический эффект тестостерона оптимально проявляется в присутствии гормона роста. Тестостерон усиливает синтез ингибитора остеоиндукции. Поэтому наступление половой зрелости тормозит рост скелета (в длину). Эстрогены (наиболее активный гормон - 17- Мощными паракринными стимуляторами остеогенеза служат различные факторы роста (фибробластов, тромбоцитов, а также трансформирующий и инсулиноподобный). Резорбция кости стимулируется, через простагландины, такими паракринными регуляторами, как интерлейкин -1 (ИЛ-1), кахексин, лимфотоксин и γ-интерферон. Цитокины - интерлейкин-1 и Но решающей остаётся регуляция с помощью кальцитонина, кальцитриола и паратгормона. Паратгормон способен осуществлять в организме следующие эффекты, определяющие ход вышеописанных процессов:
У паратгормона существует гормональный физиологический антагонист, реципрокно влияющий на кальций-фосфатный метаболизм, это гормон С-клеток щитовидной железы – кальцитонин ( КТ). Кальцитонин – пептид из 32-х аминокислот, из которых 7 остатков на аминоконце замкнуты дисульфидной связью в кольцо. Гормон синтезируется из прокальцитонина. Кальцитониновые рецепторы находятся в остеокластах, а также в клетках почек и ЖКТ. Эффекты кальцитонина сводятся к тому, что: · подавляется резорбция костного вещества остеокластами (при длительном действии нарушается остегенез остеобластами); · подавляется реабсорбция кальция и фосфата (а также Na+, Mg2+, K+) в почках; · возможно тормозит активацию макрофагов 24,25(ОН)2Dз - 24,25-дигидроксихолекальциферол, один из активных метаболитов холекальциферола - (витамина Dз), рассматривается в настоящее время как стероидный гормон. Образуется в митохондриях клеток почек, костей, хрящей, тонкого кишечника и плаценты при гидроксилировании метаболита 25(ОН) Dз по 24 атому углерода. В хрящевой зоне роста эпифизов стимулирует пролиферацию и дифференцировку хондрогенных клеток, содержащих специфические рецепторы для данного соединения. По некоторым данным, 24,25(ОН)2Dзстимулирует синтез коллагена остеобластами. Характер биологического эффекта зависит от природы клетки-мишени:
· в зрелых остеобластах и остеоцитах - уменьшение активности ЩФ, синтеза коллагена, усиление синтеза ОК и продукции цитрата; · в моноцитах - стимуляция дифференцировки в макрофаги и остеокласты, активация синтеза лизосомальных ферментов;
В почках: в проксимальных канальцах происходит стимуляция реабсорбции Р043-, а в дистальных - Са2+Механизмы процессов аналогичны тем, которые реализуются в энтероцитах. Витамин А (ретинол) - непосредственно, а также, в большей мере, в форме метаболита - ретиноевой кислоты (гормона) активирует пролиферацию и дифференцировку хондрогенных клеток в хрящевой зоне роста эпифизов, синтез хондроитинсульфатов протеогликанов; усиливает задержку SO42-, Са2+, Р043-. Действие витаминов С и К1 связано с созреванием коллагена, и Механизмы регуляции анаболических и катаболических путей тесно взаимосвязаны и призваны сохранить баланс и стабильность обмена как органических веществ, в первую очередь - белков, так и минеральных компонентов, обеспечивая константы [Са2+] и [Р043-] в крови. Гипофиз, щитовидная железа и половые железы оказывают глубокое влияние на обмен веществ и формирование костной системы. При выпадении функции гипофиза или щитовидной железы тормозится рост и развитие, нарушаются процессы дифференциации, что ведет к карликовости. Введение гипофизарных гормонов усиливает отложение кальция.
|

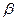 -эстрадиол-Е2) обладают общим анаболическим действием. В остеобластах, обладающих специфическими рецепторами к Е2, возрастает синтез КЛ1, активность ЩФ и, в меньшей степени, повышается синтез остеопонтина. Особенность действия Е2, по сравнению с андрогенами, заключается в чрезвычайно активной стимуляции дифференцировки хондрогенных клеток в хондроциты в хрящевой зоне роста. Данный феномен обуславливает закрытие ростовых зон, раннюю минерализацию и интраскелетную оссификацию. Следовательно, в период полового созревания происходит остановка роста скелета.
-эстрадиол-Е2) обладают общим анаболическим действием. В остеобластах, обладающих специфическими рецепторами к Е2, возрастает синтез КЛ1, активность ЩФ и, в меньшей степени, повышается синтез остеопонтина. Особенность действия Е2, по сравнению с андрогенами, заключается в чрезвычайно активной стимуляции дифференцировки хондрогенных клеток в хондроциты в хрящевой зоне роста. Данный феномен обуславливает закрытие ростовых зон, раннюю минерализацию и интраскелетную оссификацию. Следовательно, в период полового созревания происходит остановка роста скелета.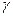 -интерферон (
-интерферон (


