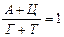Правила Чаргафа.
Аналіз нуклеотидного складу ДНК і РНК (співвідношень пуринових і пиримидинових основ) дозволив Е. Чаргафу у 1950 р. сформулювати ряд правил: 1. В ДНК молярна сума Г і А (пуринові основи) дорівнює молярній сумі Ц і Т (піримідинові основи). Ця закономірність не властива РНК, де відношення пуринових і піримідинових основ змінюється в широких межах. А + Г = Ц + Т або 2. У молекулах ДНК число залишків А завжди дорівнює числу залишків Т. В такому ж відношенні знаходяться Г і Ц. У молекулах РНК цього немає, хоча у багатьох випадках молярні співвідношення основ близькі (тут мається на увазі, що Т і У в молекулах ДНК і РНК рівноцінні). А = Т та Г = Ц,відповідно 3. Відношення суми молярних концентрацій Г і Ц до суми молярних концентрацій А і Т в ДНК і А і У в РНК 4. Кількість основ, що містять аміногрупу в положеннях 4 піримідинового та 6 пуринового ядер, дорівнює кількості основ, що містять у цих же положеннях оксогрупу. Це означає, що кількість аденіну та цитозину рівна кількості гуаніну та тиміну: А + Ц = Г + Т або Правила Чаргаффа на РНК не поширюються, оскільки РНК містить велику кількість мінорних основ. Дві перші закономірності в будові ДНК і РНК пов'язують з особливостями їх вторинної структури. У створенні вторинної структури полінуклеотидного ланцюга виняткове значення мають водневі зв'язки, що виникають між парами основ: А і Т (у РНК — А і В) і Г і Ц (у обох видах нуклеїнових кислот). Основи, створюючи пари, в яких вони поєднуються водневими зв'язками, називаються комплементарними. Характер водневих зв'язків між комплементарними основами показаний на рис. 66. З одного боку, водневі зв'язки замикаються між аміногрупами і кетогрупами, що займають певне положення в молекулах пуринових і піримідинових основ. У цьому і полягає комплементарність, тобто доповнюваність в хімічній будові пуринових і піримідинових основ: там, де у пуринової основи розташована аміногрупа, у піримідинової знаходиться кетогрупа, і навпаки. З іншого боку, водневі зв'язки утворюються за рахунок взаємодії за атомами N і NH-групам, де вони також доповнюють один одного. Саме тому в ДНК число молей А дорівнює числу молей Т, а число молей Г дорівнює числу молей Ц. Природно, що сумарний вміст пуринів і піримідинів однаковий. Третя закономірність в співвідношенні пуринових і піримідинових основ пов'язана із специфічністю ДНК і РНК. Тому відношення Г+Ц / А+Т(У) називається коефіцієнтом специфічності нуклеїнових кислот. Встановлено, що ДНК має чітко виражену видову специфічність. Особливо різкі видові відмінності характерні для ДНК. виділених з мікроорганізмів. В РНК видова специфічність відношенням виражена слабкіше. Загалом, коефіцієнт специфічності в ДНК варіює: § від 0,45 до 2,57 у мікроорганізмів, § від 0,58 до 0,94 у вищих рослин, § від 0,54 до 0,81 у тварин. У тотальної РНК коефіцієнт специфічності, як правило, вище за одиницю.
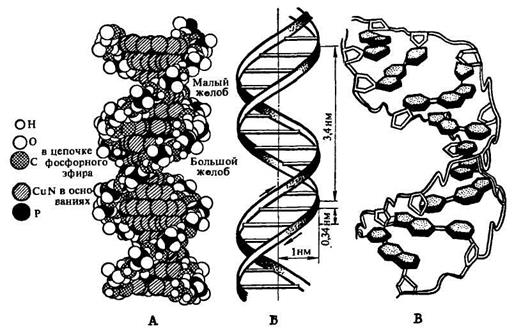
Фотографія об'ємної моделі молекули ДНК представлена на рис. 67, А. На рис. 67, Б приведена схема розташування полінуклеотидних ланцюгів і характерні їх параметри, а на рис. 67, В – схема розташування комплементарних основ, що зв'язують водневими зв'язками полідезоксирибонуклеотидні ланцюги один з одним.
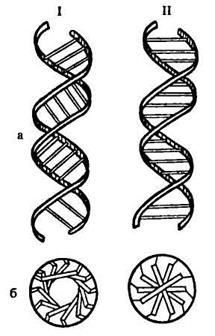
Рис. 67. Будова молекули ДНК: А — модель молекули ДНК: темні і світлі ланцюжки атомів, що виділяються на рисунку, — обернений назовні пентозофосфатний ланцюг. Всередину моделі, перпендикулярно її довгій осі, направлені пуринові і піримідинові основи, що з'єднуються водневими зв'язками: атоми, що належать основам (заштриховано криво), щільно заповнюють центральну частину моделі; Б — просторове розташування полідезоксирибонуклеотидних ланцюгів в молекулі ДНК; поперечні лінії на схемі позначають площини, в яких розташовуються пари основ, що зв'язують ланцюги один з одним: 0,34 нм — відстань між залишками сусідніх дезоксирибонуклеотидів; 1 нм — радіус молекули; 3,4 нм — крок подвійної спіралі; В — розташування комплементарних пуринових і піримідинових основ в молекулі ДНК; молекули дезоксирибози представлені білими п'ятикутниками, обведеними подвійною лінією; фосфатні групи — вигином подвійної лінії, що зв'язує залишки дезоксирибози; від кожного залишку вуглеводу відходять основи у вигляді вкритих точковим штрихуванням шестикутників і п'ятикутників: короткі подвійні лінії, що з’єднують заштриховані пуринові і піримідинові основи, представляють водневі зв'язки. Всі спіралі — правообертаючі На рис. 67 представлені моделі так званої В-форми ДНК. Річ у тому, що залежно від ряду умов ДНК існує у вигляді різноманітних впорядкованих волокнисто-кристалічних структур. Їх отримано більше десяти, чотири з них: А-, В-, С- і Т-форми ДНК — вивчені методом рентгеноструктурного аналізу. При дотриманні загального плану будови молекули ДНК у вигляді біспірального полідезоксирибонуклеотиду вони відрізняються за рядом параметрів, властивих кожній з названих конформаційних модифікацій ДНК (таблиця. 20). Найбільш сильна зміна конформації відбувається при перетворенні А-форми ДНК на В-форму. Зокрема, при цьому різко зміщується просторова структура β-D-2-дезоксирибози у її складі, внаслідок чого кут нахилу площини основ до осі спіралі змінюється більш ніж на 20° (рис. 68).
У біологічному сенсі В-форма найбільш адекватна для реплікаційних процесів, А-форма — для процесу транскрипції, С-форма — для упаковки ДНК у складі надмолекулярних структур хроматину і деяких вірусів. Таким чином, вторинна структура молекул ДНК пов'язана із здійсненням інформаційних процесів в живій природі, а саме: § А-форма ДНК — з передачею інформації від ДНК до РНК, § В-форма ДНК — з множенням кількості інформації, § С-форма ДНК — із зберіганням інформації.
Таблиця 20. Характеристика деяких конформаційних станів ДНК
Два види сил утримують два полідезоксирибонуклеотидних ланцюги в біспіральній молекулі ДНК: По-перше, це водневі зв'язки між комплементарними азотистими основами, спрямованими всередину подвійної спіралі ДНК (див. рис. 66). Утворюючи водневі зв'язки, основи знаходяться в площині, перпендикулярній подовжній осі В-форми ДНК, так що ці зв'язки діють у поперечному напрямі. По-друге, це сили гідрофобних взаємодій між азотистими основами, зібраними в «стопку» уздовж молекули ДНК; при такій упаковці основ у водному середовищі виникають сили, що перешкоджають контактам неполярних (гідрофобних) основ з молекулами води, внаслідок чого основи зближуються, а стопкоподібна упаковка зміцнюється внаслідок їх міжплощинних взаємодій одна з одною. Їх називають тому стекінг-взаємодіями (тобто взаємодіями, спрямованими уздовж стопки основ і, в результаті, вздовж молекули ДНК). Останнім часом силам стекінг-взаємодії надають істотніше, ніж раніше, значення в підтримці вторинної структури ДНК, а водневим зв'язкам між комплементарними парами основ визначають загалом направляючу роль у взаємній орієнтації основ в процесі стекінг-взаємодії. Дійсно, вклад гідрофобних взаємодій в підтримку вторинної структури біспіральних полінуклеотидів зростає при заміні У на Т, тобто за рахунок гідрофобного метильного радикалу, що додатково вводиться.
|
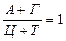
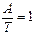 ;
; 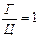 .
.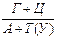 в обох видів нуклеїнових кислот сильно варіює. Особливо широкі кордони мінливості цього показника в ДНК.
в обох видів нуклеїнових кислот сильно варіює. Особливо широкі кордони мінливості цього показника в ДНК.