
Основные особенности структуры геномов прокариот.
ПЕРЕСЕЧЕНИЕ ДВУХ ПРЯМЫХ. Система имеет единственное решение:
УГОЛ МЕЖДУ ПРЯМЫМИ.
ПЕРПЕНДИКУЛЯРНЫЕ ПРЯМЫЕ.
Пример: Написать уравнение прямой, проходящей через точку A (2; –1) перпендикулярно 3 x + 2 y – 4 = 0.
ПАРАЛЛЕЛЬНЫЕ ПРЯМЫЕ.
M (3; –1) – точка пересечения диагоналей. Написать уравнение двух других сторон.
Решение: 1) Находим точку B: 2) M – середина BD,
3) (DC)||(AB),
4) (AD)||(BC),
СОВПАДАЮЩИЕ ПРЯМЫЕ.
ВЗАИМНОЕ РАСПОЛОЖЕНИЕ ДВУХ ПРЯМЫХ.
ПЕРЕСЕЧЕНИЕ ДВУХ ПРЯМЫХ. Система имеет единственное решение:
УГОЛ МЕЖДУ ПРЯМЫМИ.
ПЕРПЕНДИКУЛЯРНЫЕ ПРЯМЫЕ.
Пример: Написать уравнение прямой, проходящей через точку A (2; –1) перпендикулярно 3 x + 2 y – 4 = 0.
ПАРАЛЛЕЛЬНЫЕ ПРЯМЫЕ.
M (3; –1) – точка пересечения диагоналей. Написать уравнение двух других сторон.
Решение: 1) Находим точку B: 2) M – середина BD,
3) (DC)||(AB),
4) (AD)||(BC),
СОВПАДАЮЩИЕ ПРЯМЫЕ.
Вопрос 18. Основные особенности структуры геномов прокариот. Геном – это совокупность всех генов организма с регуляторными элементами и все молчащие участки. У прокариот и эукариот геномы отличаются. Величина генома у бактерий варьирует в пределах от 0,8 до 8×106 пар оснований. В бактериальной клетке – 2 типа молекул ДНК: 1) хромосома (нуклеоид) Это самая крупная молекула, всегда кольцевая, длина – несколько миллионов пар нуклеотидов (мегабаз). В молекуле расположен весь набор генов. Молекула имеет 1 точку инициации репликации (ori), т.е. молекула самореплицируется. 2) плазмида (может отсутствовать) Несет в себе, например, гены устойчивости к антибиотикам. Также содержит 1 ori, самореплицируется. Ori хромосомы и плазмиды совпадают, т.к. разводить вторую систему репликации экономически не выгодно. Важнейшая особенность в организации бактериального генома состоит в его разделении на два во многих отношениях симметричных полугенома. Так, у E.coli и Salmonella typhimurium область начала двунаправленной репликации – oriC – разделяет хромосому на два примерно равных по величине плеча, отделенных на противоположной стороне кольцевой хромосомы областью терминации репликации – terC.
ДНК бывает: 1) кодирующая (содержит «гены», гены — это участки ДНК, несущие какую-либо целостную информацию — о строении одной молекулы белка или одной молекулы РНК.) 2) некодирующая (содержит регуляторные элементы, обеспечивающие процессы репликации, транскрипции). По сути некодирующая ДНК – это промежутки между генами и регуляторными элементами, между регуляторными элементами. Промежутки между регуляторными элементами – спейсеры. Спейсеры бывают транскрибируемые (они находятся внутри оперонов между рамками считывания) и нетранскрибируемые (они находятся между оперонами). В транскрибируемых спейсерах есть участки, которые играют роль для трансляции, нетранскрибируемые участки – не нужны бактериям.
Бактериальный геном очень компактен, т.к. только 1 ori и кольцо должно быть маленьким. Некодирующая часть невелика (не более 300 пар оснований). Для хромосом бактерий в отличие от хромосом эукариот характерно относительно высокое содержание генов в расчете на имеющуюся ДНК.
Кодирующая часть ДНК состоит из открытых рамок считывания (ORF). ORF – это последовательность нуклеотидов, кодирующих белки. Последовательность ДНК от точки начала транскрипции до точки терминации – транскриптон. У бактерий между точками инициации и терминации транскрипции имеется несколько ORF. В сумме все выше перечисленное называется – оперон (это бактериальный транскриптон, состоящий из нескольких ORF). Наличии оперонов – целостно транскрибируемых групп функционально родственных генов – еще 1 особенность прокариотического генома.
Т.к. геном бактерий состоит из ORF, то можно сделать вывод, что у бактерий генов нет. Рамки считывания многообразные, они кодируют все белки, т-РНК, р-РНК, дополнительные структуры (их кодируют гены домашнего хозяйства, эти гены по составу примерно одинаковые).
Существуют участки инициации транскрипции – промоторы. Это ограниченная область, с которой связывается РНК-полимераза. В промоторе есть 3 функциональные зоны: 1) домен Прибного (ТАТААТ) 2) домен -35 (ТТGАСА) 3) старт транскрипции
Стоит отметить, что процессы регуляции транскрипции у эукариот и прокариот различаются также по причине различий в их геномах. Так, регуляция на уровне трансляции характерна только для прокариот и связана с наличием аттенюаторов (ослабителей). Аттенюаторы могут быть только в бактериях, т.к. у эукариот процессы транскрипции и трансляции разделены в пространстве. Аттенюаторы – это регулируемые терминаторы бактерий. Впервые обнаружен и лучше других изучен аттенюатор триптофанового оперона E. coli. Аттенюатор расположен на расстоянии 180 п.н. от стартовой точки транскрипции. Перед аттенюатором располагается несколько областей с центральной симметрией. Образующаяся на них лидерная РНК содержит 4 участка, способных образовывать шпильки. Могут образовываться шпильки 1:2 и 3:4, либо образуется шпилька 2:3. Шпилька 3:4 – терминаторная шпилька.
Также регуляция транскрипции осуществляется на уровне инициации. Например, это регуляция лактозного оперона или система сАМP-зависимой регуляции катаболитных оперонов. В случае лактозного оперона с регуляторной областью связывается репрессор, который ослабляет связь РНК-полимеразы и промотора. Лактоза выступает в качестве индуктора. Она связывается с репрессором, в результате он не может связаться с оператором, промотор активируется и идет синтез ферментов, перерабатывающих лактозу. В случае сАМP-зависимой регуляции катаболитных оперонов с регуляторной областью связывается белок-активатор. Его связывание упрочняет комплекс РНК-полимеразы с промотором.
|


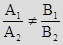


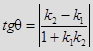


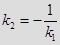


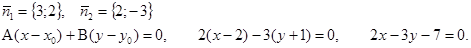

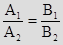

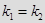
 Пример:
Пример: – стороны параллелограмма;
– стороны параллелограмма;
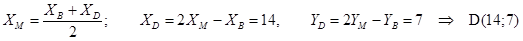




 – это и есть условие совпадения двух прямых.
– это и есть условие совпадения двух прямых.



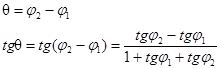











 Пример:
Пример: – стороны параллелограмма;
– стороны параллелограмма;

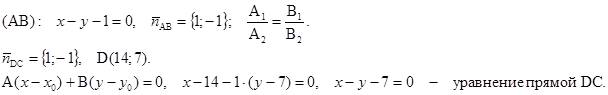



 – это и есть условие совпадения двух прямых.
– это и есть условие совпадения двух прямых.




