КОЛЕБАНИЯ В СИСТЕМЕ «ХИЩНИК – ЖЕРТВА». Математическая модель взаимоотношений хищника и жертвы.
Еще в 20-х годах 20 века А. Лотка (1925), а позднее В. Вольтерра (1926) предложили математические модели, описыващие сопряженные колебания численности популяций хищника и жертвы.
В лабораторных условиях одним из первых примеров колебания численности хищника и жертвы был получен Г.Ф. Гаузе (1934). Объекты – инфузории Paramecium caudatum (жертва) и Didinum nasutum (хищник). Пищей для Paracium caudatum служили регулярно вносимые в среду бактерии. Данная система оказалась крайне неустойчивой: пресс хищника по мере увеличения его численности привёл к полному истреблению жертвы, а потом и самого хищника (рис. 5). При устройстве убежищ для жертвы (например, в виде стеклянной ваты) часть особей жертвы могла в них спрятаться и оказаться недоступной для хищника. В результате все особи хищника погибали, а численность особей жертвы восстанавливалась за счет размножения особей, уцелевших в убежище (рис. 6). Некоторое подобие колебаний численности хищника и жертвы Г.Ф. Гаузе удалось добиться в том случае, когда он время от времени вносил в культуру и хищника, и жертву, иммитируя таким образом иммиграцию (рис. 7). Сосуществование конкурирующих видов и отношения типа «хищник—жертва» часто усиливаются во фрагментированных местообитаниях, поскольку более слабые конкуренты или жертвы получают определенные преимущества. Опыты К. Хаффейкера. Классическим примером служит эксперимент, в котором были задействованы два вида клещей (растительноядный и хищник), несколько апельсинов (пища для растительноядного клеща), резиновые мячи и банка вазелина. В относительно однородной среде (апельсины, расположенные близко друг к другу) хищники вскоре находили и поедали всю добычу, а затем умирали голодной смертью. Однако, если среду делали фрагментированной, перемежая апельсины резиновыми мячами, и затрудняли пути хищникам, создавая вазелиновые барьеры, хищники и жертвы вполне сносно сосуществовали. Жертвы находили относительно безопасные места. В любой отдельный промежуток времени система состояла из свободных участков, участков, занятых видом-жертвой, и участков, в которых хищники ловили жертв. В природных условиях циклические изменения численности двух видов, связанных пищевыми отношениями, выявить труднее, т.к. на их размножение и гибель влияют взаимодействия со многими другими видами, а также абиотические условия среды. Однако примеры колебаний численности в системе «хищник-жертва» можно найти и в природе, особенно в обедненных сообществах, где межвидовые связи не столь разнообразны. Бабочка кактусовая огневка (Cactoblastis cactorum), завезенная в Австралию, значительно снизила численность кактуса опунции (но не уничтожила его полностью потому, что кактус успевает расселиться немного быстрее). Обычно эти бабочки откладывают яйца на некоторые растения, показавшиеся им в силу каких-то причин особо привлекательными. Вылупившиеся личинки при высокой плотности обычно полностью уничтожают свое кормовое растение. В тех местах, где опунция истребляется полностью, перестает встречаться и огневка. Если сюда вновь проникнет опунция, в течение времени ее растения произрастают без риска быть уничтоженными огневкой. Со временем, однако, огневка снова появляется и уничтожает опунцию. Другие примеры: 9-10 - летние циклы колебания численности зайца и рыси, 3-4-летние циклы колебания численности в тундровой зоне леммингов и их хищников – песцов и полярных сов. Однако колебания численности в системе «хищник-жертва» не обязательно являются подтверждением модели Лотки-Вольтерры. Так, если численность популяции растительноядного животного колеблется, то это может отражать как её взаимодействия с пищевыми объектами, так и с хищником. Таким образом, взаимодействия типа «хищник-жертва» могут порождать регулярные колебания численности взаимодействующих популяций и могут усиливать такие колебания, если они существуют вследствие других причин. Есть данные о том, что на ряде островов, где рыси и других истинных хищников нет, численность зайца также подвержена циклическим колебаниям. Существует предположение, что это связано с изменением количества и качества растительного корма для зайцев (они потребляют кончики побегов кустарников и небольших деревьев). Сильное объедание побегов приводит к образованию новых побегов с высоким содержанием ядовитых для зайца веществ, что делает их несъедобными. Растения остаются защищенными от поедания в течение двух-трех лет после сильного обгрызания их зайцами. Таким образом, видимо, данный пример свидетельствует о существовании связи между количеством и качеством растительного корма и численностью популяции зайца. Колебания же численности популяции хищника здесь скорее всего следуют за колебаниями численности популяции зайца, а не вызывают их. Однако здесь хищники, вероятно, способствуют тому, что эти колебания выражены более отчетливо. Отношения хищник-жертва обеспечивают перенос энергии с одного трофического уровня на другой и обеспечивают устойчивость сообщества. Благодаря деятельности хищников регулируется численность жертв, это позволяет выжить и другим видам этого трофического уровня, в результате увеличивается биоразнообразие сообществ.
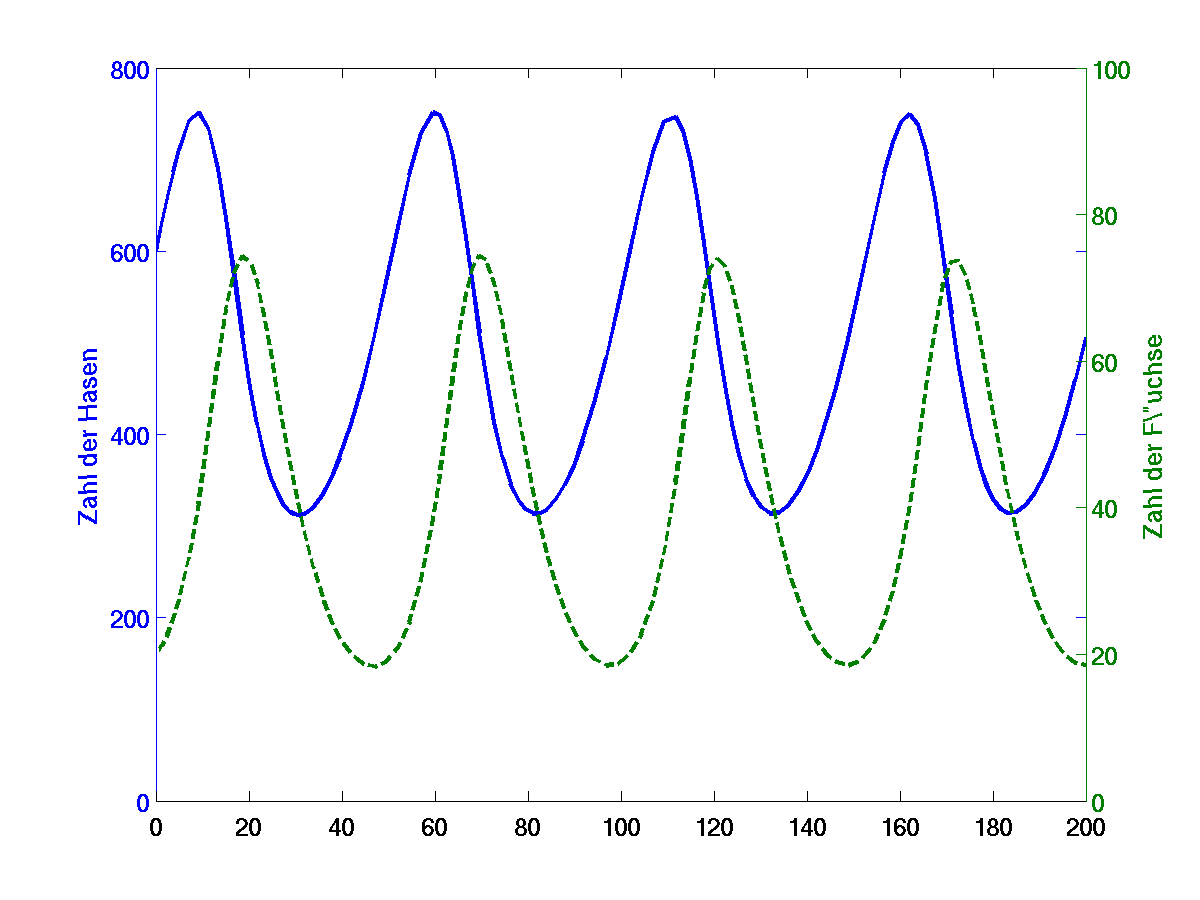
Рисунок 4.
|
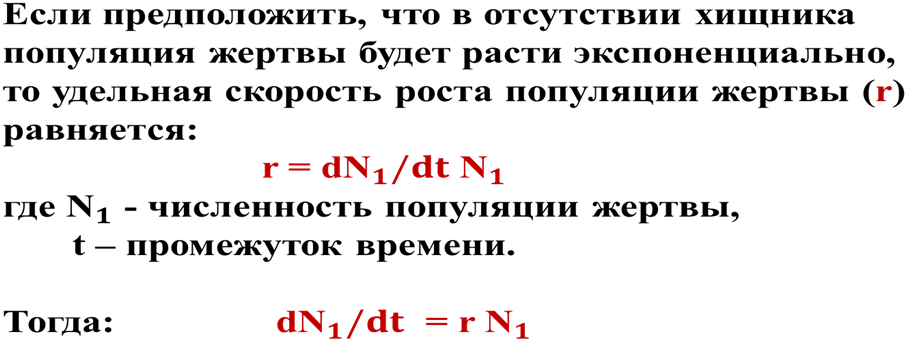
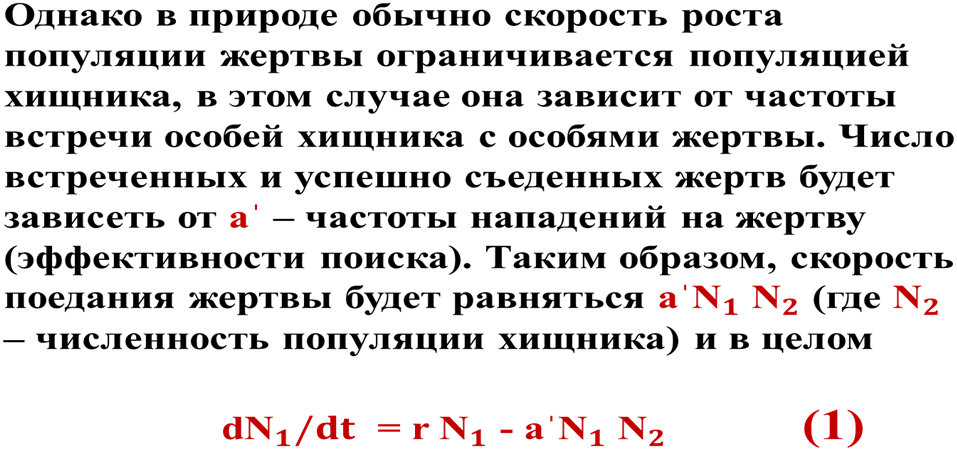
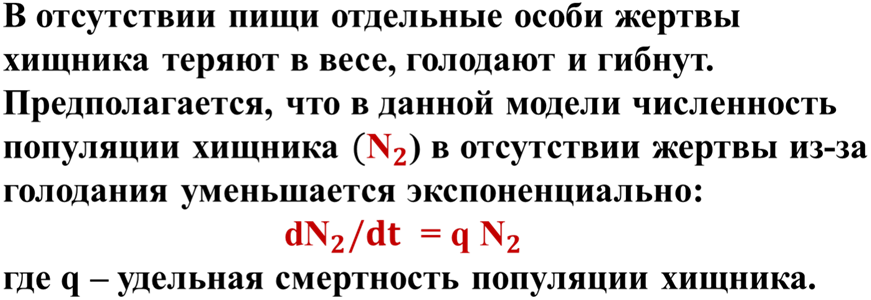
 Согласно этой модели в системе «хищник-жертва» происходят неограниченные сопряженные колебания численности популяций хищника и жертвы (т.е. оба вида взаимно контролируют численность друг друга): когда велико число жертв, численность хищника увеличивается, что приводит к повышению пресса хищников на популяцию жертвы и тем самым к снижению ее численности. Это снижение в свою очередь ведет к ограничению хищников в пище и к падению их численности, которое вызывает ослабление пресса хищников и увеличение численности жертвы, что снова приводит к росту популяции хищника и т.д. (рис. 4).
Согласно этой модели в системе «хищник-жертва» происходят неограниченные сопряженные колебания численности популяций хищника и жертвы (т.е. оба вида взаимно контролируют численность друг друга): когда велико число жертв, численность хищника увеличивается, что приводит к повышению пресса хищников на популяцию жертвы и тем самым к снижению ее численности. Это снижение в свою очередь ведет к ограничению хищников в пище и к падению их численности, которое вызывает ослабление пресса хищников и увеличение численности жертвы, что снова приводит к росту популяции хищника и т.д. (рис. 4).