
Елементарна ландшафтно-екологічна територіальна одиниця – геотоп.
Критерії виділення геотопу. Ніякої природної межі (ґрунтової, геоботанічної тощо) через геотоп провести не можна, вони можуть лише збігатися з його межами. Виходячи з цього, фація (геотоп) є об'єктивним природно однорідним підрозділом ландшафту, який сама природа наділила територіальною неподільністю. Більш наближений до реальності підхід до виділення геотопів пов'язаний з розчленуванням території на однорідні ділянки за кожним з геокомпонентів та факторів топічної диференціації ландшафту. У німецькій геоекологічній літературі такі ділянки називаються топами. Виділяються морфотопи (однорідні елементарні поверхні рельєфу), літотопи (ділянки, однорідні за геологічною будовою), гідрогопи (за умовами зволоження), кліматопи (за мікрокліматом), педотопи (за грунтом), фітотопи (за рослинністю), зоотопи (за тваринним населенням). їх просторовий збіг та взаємодія утворюють комплексну територіальну одиницю— геотоп. 1.Основою виділення топів є градієнтний критерій. Виділені за градієнтним критерієм топи можуть бути гомогенними (незначна варіація значень характеристик у межах топів) або ж клінальними (значення змінюються в певному напрямку, але різких змін в межах топу немає). Клінальними часто є педотопи прямих схилів, де вниз по схилу зменшується потужність грунту. Переважна більшість фітотопів також клінального типу, що пояснюється континуальним характером рослинного покриву
2. Статистичний критерій виділення геотопу зводиться до того, що варіація значень змінних у його межах має бути меншою, ніж ця варіація між різними геотопами (внутрішньогрупова дисперсія менша за міжгрупову). 3. За географічним критерієм можна відрізнити геотопи від деяких інших природних територіальних об'єктів — мурашника, ями, утвореної поваленим деревом, а також антропогенних — дороги; каналу тощо. За такий критерій можна прийняти той, що запропонував О. І. Перельман (1960): геотоп не має внутрішніх властивостей, які унеможливлювали б його теоретично необмежене збільшення площі. Так, невелика пляма солончаку теоретично може (і в природних ландшафтах це не рідкість) розростися до значних розмірів, а для ями з-під поваленого дерева, мурашника, дороги це неможливо. 4. Критерій відносної статичності. Ділянки, які на загальному просторовому фоні виділяються внаслідок дії тимчасових (внутрішньорічних, річних ритмічних або флуктуаційних) факторів, геотопами не вважаються. Геоботанічні аспекти (ділянки фітоценозу з домінуванням певного коротковегетуючого виду), ділянки перезволоженого грунту після дощу, ділянки надвипасу, що заростають за кілька вегетативних періодів, та подібні короткочасні територіальні утворення є елементами внутрішньотопічної структури. 5. За картографічним критерієм геотопи виділяють таких розмірів, які можна показати на крупномасштабній карті і які ще можна розглядати як територіальні операційні одиниці при вирішенні практичних завдань. Щодо мінімального розміру геотопу висловлено кілька пропозицій. Т. Бартковскі (1971) вказує 100 м2 як критичну площу ділянки, менше якої виділяти геотопи немає рації. Ван-Леєвен (1981) за такий розмір вважає 500 м2, а Р. Форман та М. Годрон (1986) називають ще більші величини — шириною від 10 до 1000 м.
Внутрішньотопічна територіальна структура. Територіальна однорідність геотопу відносна, а тому його можна поділити на ще більш однорідні частини, тобто він має внутрішньотопічну територіальну структуру. Здебільшого основним геокомпонентом, що визначає або індикує внутрішньотопічну структуру геотопу, є рослинність. Ієрархічні рівні внутрішньотопічної диференціації: Перший рівень визначається дією на середовище окремих особин рослин, другий — територіальними особливостями їх розмноження, третій, пов'язаний із взаємодією близькорозташованих рослин. Італійський еколог Г. Негрі у 1954 р. запропонував термін екоїд. Розмір екоїдів, утворених деревами, трохи перевищує проекцію їх крони, а дерновинними злаками — менший за проекцію їх кореневої системи. Екоїди окремих рослин можуть перетинатися і, навпаки, у межах геотопу можуть «бути ділянки, де екоїдів немає зовсім. Другий ієрархічний рівень територіальної структури фітотопу пов¢язаний з особливостями розмноження рослин. Насамперед -вегетативне розмноження, яке призводить навіть до виникнення ділянок одновидових заростей (наприклад, чорниці). Такі ділянки Т. А. Работнов (1972) називає клоновими. Вони зустрічаються далеко не в усіх геотопах. Територіальні елементи геотопу третього рів'ня утворюються в разі тісних зв'язків між окремими особинами рослин. Ці зв'язки проявляються насамперед у конкуренції за місцеві ресурси (світло, вологу, поживні речовини), алелопатичних взаємодіях. Вони можливі лише за досить близького взаєморозташування особин, тобто при перекритті їх екоїдів. Такі структурні одиниці геотопу:можна назвати поліекоїдами З ландшафтно-екологічної точки зору до трьох рівнів територіальної
3. Відношення між геотопами та типи ландшафтних територіальних структур. Поняття про ландшафтну територіальну структуру. Будь-яка геосистема, рангом вища за геотоп має певну ландшафтну територіальну структуру. Елементами цієї структури є геосистемн нижчого рангу, ніж досліджувана Ними можуть бути геотопи, які за визначеним типом та характером просторових зв'язків між ними обєднуються у ландшафтні територіальні одиниці вищих рангів. Сукупність цих одийиць становить таксономічний ряд геосистем, а їх взаємне розташування в межах певної геосистемн визначає і конфігураційні та топологічні особливості (площу, форму, складність структури). Таким чином, ландшафтну структуру можна визначити як сукупність ландшафтних територіальних одиниць, конфігураційно та ієрархічно впорядкованих просторовими відношеннями певного типу. Тип відношень між геотопами є основою виділення відповідного типу ландшафтної територіальної структури (ЛТС), оскільки визначає: принцип інтеграції геотопів у більш складні ландшафтні територіальні одиниці; таксономічний ряд останніх (ієрархічну впорядкованість ЛТС); ареали, межі, взаєморозташування однорангових ландшафтних одиниць (конфігураційну впорядкованість ЛТС). Щоб виділити ЛТС певної території, необхідно: 1) задати тип просторових відношень між геотопами; 2) виявити множини геотопів, закономірно зв'язаних цими відношеннями; 3) для кожної з цих множин здійснити інтеграцію геотопів у ландшафтні територіальні одиниці таксономічного ряду ЛТС, що виділяється; 4) встановити та показати на карті ареали, межі, взаєморозміщення ландшафтних територіальних одиниць одного рангу. Згідно з принципом ландшафтної поліструктурності, у певному регіоні можна виділити ЛТС різних типів і відповідно скласти ландшафтні карти, кожна з яких моделювала б певний аспект (структуру) територіальної організації ландшафту. Основні типи ландшафтних територіальних структур. Оскільки геотопи пов'язані між собою різними просторовими відношеннями, вони можуть інтегруватися в різні ЛТС залежно від того, яке структуроформуюче відношення прийнято за основу цієї інтеграції. З багатьох просторових відношень між геотопами виділяються п'ять їх типів, які разом визначають основні риси ландшафтної територіальної організації в цілому та можливість вирішення переважної більшості практичних завдань ландшафтної екології. Цими відношеннями та відповідними їм типами ЛТС є: 1. Генетико-морфологічні — відношення спільності походження (генезису) та еволюції геотопів, які знаходять вираз у їх будові (морфології) і формують генетико-морфологічний" тип ЛТС; 2. Позиційно-динамічні— зв'язок геотопів горизонтальними речовинно-енергетичними потоками та їх відношення до ліній зміни інтенсивності цих потоків; парагенетичні - відношення геотопів до ліній концентрації горизонтальних потоків; 3. Басейнові ландшафтні — спільність геотопів за гідрофункціонуванням та їх відношення до басейнів по-
верхневого стоку; біоцентрично-сітьові — біотичні міграції організмів та окремих популяцій між геотопами. Кожний тип ЛТС має власний таксономічний ряд ландшафтних територіальних одиниць.
Тема: V Ландшафтні територіальні структури (хорологічна ландшафтна екологія) 1. Генетико-морфологічна ландшафтна територіальна структура. 2 Позиційно-динамічна ландшафтна територіальна структура. 3. Парагенетична ландшафтна територіальна структура. 1. Генетико-морфологічна ландшафтна територіальна структура. Структуроформуючі відношення — генетико-морфологічна однорідність геосистем. Основою виділення одиниць генетико-морфологічної ЛТС є об'єднання територіально суміжних геотопів у більші геосистеми за принципом спільності їх походження (генезису), часу виникнення та закономірностей розвитку (еволюції).. Близькі в генетико-еволюційному відношенні геосистеми мають і однотипну геокомпонентну будову Таким чином, морфологічні ознаки як внутрішньої будови (складу геокомпонентів), так і зовнішнього вигляду геосистем є важливим індикатором їх генетико-еволюційиих особливостей і тому розглядаються як критерії групування морфологічно подібних геотопів в одиниці генетико-морфологічної ЛТС. Г енетико-морфологічна однорідність геосистем - це представленість у геосистемі лише генетично близьких таксонів усіх її геокомпонентів (повна однорідність) або деяких з них (часткова однорідність, наприклад — геолого-геоморфологічна, ґрунтова, фітоценотична тощо). Діагностичними геокомпонентами геосистем регіонального та нижчих рівнів розмірностей (тобто компонентами, за якими виділяються геосистеми) є геологічна будова, грунти, рослинність, грунтові води, а також рельєф як важливий фактор просторової диференціації ландшафтів. Рівні генетико-морфологічної однорідності території за її різними діагностичними геокомпонентами. Геологічна будова: I рівень однорідності (Ь —/)—однорідною є територія, формована па одному геологічному фундаменті 2-ий рівень і / _2) — територія, однорідна за складом дочетвертинних порід, 3-й рівень (Ь — 3) —територія, верхні верстви геологічної будови якої близькі за літолого-генетичними характеристиками (наприклад, складена елювіальними, делювіальними, лесовими суглинками) 4-ий півень (Ь — 4) —ділянка з одним типом поверхневих відкладів, допускається їх різна потужність. 5-ий рівень (Ь —5)—ділянка, зайнята одним літотопом Рельєф: 1-ий рівень однорідності (Я — 1) —територія займає комплекс мезоформ рельєфу одного генезису і віку (наприклад, структурно-денудаційні пліоценові рівнини з такими мезоформами, як гряди та міжгрядові улоговини). 2-й рівень (/?—2) — територій ''однієї мезоформи рельєфу (гряди, улоговини, річкові долини з' терасовим комплексом); допускаються морфотопи, накладені на1 мезоформу (ерозійні форми, конуси виносу тощо); 3-й рівень (Н—3) — територія, зайнята одним елементом рельєфу або його малою формою (неглибоко врізані лощини, промоїни, карстові во-ронки, западини тощо); 4-й рівень (К — 4) — елементарні поверхні (грані) рельєфу, де немає ліній перегинів схилу (територія одного морфотопу). Ґрунтовий покрив: 1-й рівень однорідності (5 — 1) —територія, рівнинні поверхні якої зайняті грунтами одного зонального типу грунтоутворюючого процесу (наприклад, дерново-степового тощо), 2-й рівень (З — 2) —територія з грунтами, послідовність змін яких у часі зумовлена різними факторами, виділяються літосерії (слабко-, середньо-, сильнозмиті чорноземи звичайні; потужні, середньопотужні, малопотужні, короткопрофільні чорноземи звичайні) та ін. Усі вони беруть початок від одного виду грунту (чорнозему звичайного середньогумусного) і характеризують його еволюцію під впливом певного фактора (змиву тощо). 3-й рівень (З — 3) —територія, переважна більшість грунтів якої формують один головний генетичний ряд, від якого можуть бути відгалуження інших рядів, зумовлених факторами, що супроводжують основний; здебільшого грунти належать до одного генетичного підтипу. 4-й рівень (3—4) —ділянка, зайнята грунтами одного генетичного ряду, можливі контрастні межі між ними. 5-й рівень (5 —5) — ділянка, зайнята однією ґрунтовою відміною або кількома відмінами одного генетичного ряду за відсутності різких меж між ними (один педотоп). Рослинний покрив: 1-й рівень (Р —/) —територія з домінуванням одного класу рослинних формацій зонального типу; допускається широкий діапазон інтразональних асоціацій. 2-й рівень (Р — 2) — територія з рослинними угрупованнями, послідовності змін яких у часі зумовлені різними факторами (такі послідовності називаються сукцесійними рядами), сходяться до однієї спільної клімаксової асоціації, тобто утворюють моноклімаксову сукцесійну систему. 3-й рівень (Р — 1) —територія, більшість асоціацій якої належать до одного сукцесійного ряду, а деякі утворюють відгалуження від нього; можуть бути асоціації різних рослинних формацій. 4-й рівень (Р — 4) —ділянка, асоціації якої належать до одного сукцесійного ряду, переважно однієї формації; можливі контрастні межі між фітотопами. 5-й рівень (Р —5) — ділянка виділу асоціації або клінального фітоценозу (один фітотоп). Грунтові води: 1-й рівень (О№—/) —територія, грунтові води якої знаходяться в одному водоносному горизонті; допускається широка просторова амплітуда глибин залягання ґрунтових вод, різні типи їх хімізму та ступені мінералізації. 2-й рівень (СРР-— — -2) — територія з гідротопами одного типу водного режиму (промивного, випітного тощо — див. § 3.2), допускається варіація хімізму, ступеня мінералізації ґрунтових вод, 3-й рівень (СШ — 3) — територія з одним типом хімізму, ступенем мінералізації та рівнем ґрунтових вод (один гідротоп). Відповідно названим рівням територіальної однорідності геокомпонентіввиділяються і геосистеми хоричної розмірності різних рангів. Територіальні одиниці. Елементарною одиницею генетико-мор-фологічної ЛТС є геотоп. Послідовно об'єднавши суміжні геотопи у більші за розміром територіальні одиниці так, що вони стають генетико-морфологічно однорідними відносно певного рівня, можна виділити геосистеми чотирьох таксономічних рівнів: наногеохора — мікрогеохора — мезогеохора — макрогеохора. Наногеохора сукупність суміжних геотопів, розташованих па одній елементарній поверхні рельєфу (в одному морфотопі), геологічна будова якої відрізняється лише потужністю верхніх грунтоутворюючих верств порід, водний режим одного типу, грунти одного генетичного ряду, рослинні угруповання одного сукцесійпого ряду, причому діапазон видів грунтів та рослин них асоціацій не виходить за межі генетичного підтипу (грунти) та формації (рослинність). Розміри наногеохор: в ерозійно почленованих ландшафтах становлять 0,2—0,5 км2, плоских рівнинах з глибоким рівнем залягання ґрунтових вод 0,5—2,0, в горах та на низовинах із засоленими грунтами 0,1—0,3 км2 і менше. До наногеохори близьке, але трохи менше за об'ємом підурочище класичного ландшафтознавства. Мікрогеохора — сукупність суміжних наногеохор, розташованих на одному елементі рельєфу або одній його малій формі, геологічна будова яких неоднакова лише за потужністю літолого-генетично близьких поверхневих відкладів, різниця в рівні ґрунтових вод не приводить до формування різних типів водного режиму, грунти формують основний генетичний ряд, від окремих ланок (видів грунту) якого можуть відгалужуватися короткі ряди, зумовлені фактором, що накладається на провідний фактор диференціації та динаміки педотопів, рослинні угруповання утворюють один головний сукцесійний ряд з короткими відгалуженнями від нього. Розміри мікрогеохор на рівнинному рельєфі з однорідним лесовим покривом при глибокому рівні ґрунтових вод (Північне Причорномор'я) площа мікрогеохор може досягати 100 км2 і навіть більше. Середні її розміри становлять 3—8 км2 на ерозійному рельєфі та 10—25 км2 на рівнинному. Мезогеохора — це територія з одним геологічним фундаментом та складом вкриваючих його дочетвертинних порід, різними генетичними типами четвертинних відкладів, розташована на одній мезоформі рельєфу спільного походження (річкова долина з терасовим комплексом, дельтовий комплекс, гряда, міжгрядова улоговина тощо), з ґрунтовими водами одного водоносного горизонту, грунтами та рослинними угрупованнями, ряди яких сходяться до одного центру (відповідно — до модального виду грунту та клімаксової асоціації). На відміну від мікрогеохор, грунтово-рослинннй покрив яких сформувався під впливом одного провідного фактора, в мезогеохорі таких факторів кілька. Мезогеохори займають досить значні площі (0,5—2,0 тис. км2) і є зручними одиницями регіонального управління та проектування, оскільки співрозмірні з адміністративно-управлінськими територіальними одиницями нижчих рангів (територіями агровиробничих підприємств, муніципалітетами, районами тощо). Макрогеохора — найбільша з геосистем хоричної розмірності та найменша з регіональних. її слід розглядати як однорідну в регіональному відношенні та закономірно різнорідну в хоричному. Закономірність у різноманітності геокомпонентів, що складають макрогеохору, полягає в обмеженості типів геотопів, які можуть у ній формувалися. Це обмеження є результатом фільтрації регіональними факторами можливих сполучень окремих геокомпонентів у вертикальній структурі геосистеми Макрогеохора займає територію, що складається з комплексу мезоформ рельєфу близького генезису та віку, утворених на одному геологічному фундаменті (одному субстраті формування рельєфу). Однорідність її грунтово-рослинного покриву визначається зональними факторами, тому на рівнинах представлені грунти одного типу грунтоутворюючого процесу та асоціації одного класу рослинних формацій.
Розміри макрогеохор коливаються від 1—3 тис. км2 (гірські регіони) до 6—10 км2 (рівнини) і їх можна зображати на дрібномасштабних картах. 2 Позиційно-динамічна ландшафтна територіальна структура. Структуроформуючі відношення. Групування геотопів у територіальні одиниці позиційно-динамічної ЛТС грунтується на їх відношенні до ландшафтних рубежів, вздовж яких змінюються інтенсивність та напрямок горизонтальних іречовинно-енергетнч-них площинних потоків. Носіями цих потоків можуть бути мобільні геокомпоненти — вода, повітря та живі організми. Разом з ними відбувається міграція й інших речовин, зокрема техногенних. забруднень. Горизонтальні потоки води в ландшафті стійкі за напрямком і здатні односпрямованими зв'язками інтегрувати геотопи в територіальні структури. З потоком води по земній поверхні переміщуються частки грунту (площинна ерозія), хімічні елементи та сполуки (в тому числі добрива, отрутохімікати), мікроорганізми, змивається насіння культурних рослин. Ці процеси охоплюють великі площі, а їх екологічні наслідки вельми суттєві. Територіальні одиниці позиційно-динамічної ЛТС виділяють так, щоб інтенсивність сучасних та потенційно можливих процесів, зумовлених горизонтальними речовинно-енергетичними потоками на них, була майже однаковою. Тому межі між ними проводять вздовж ліній стрибкоподібної зміни градієнтів горизонтальних потоків. Такі лінії можна назвати каркасними лініями динаміки ландшафту. Ними є всі каркасні лінії рельєфу— вододільна, тальвегу, підошви, бровки схилу, лінії його перегинів. Роль каркасних ліній динаміки ландшафту відіграють також межі між геотопами, що відрізняються фільтраційними властивостями грунтів та порід зони аерації. Вздовж цих меж різко змінюється інтенсивність фільтрації води та умови міграції хімічних елементів (у ландшафтно-геохімічному відношенні ці лінії відповідають латеральним механічним бар'єрам). Межі між грунтами з різною протиерозійною стійкістю також приймаютьсяза каркасні лінії динаміки ландшафту У сучасному ландшафті роль таких ліній відіграють також і деякі антропогенні лінійні елементи, вздовж яких змінюються параметри стоку (дороги на насипу, канали, лісосмуги тощо). Позиція геотопів щодо каркасних ліній динаміки ландшафту визначає їх динамічні особливості, зумовлені горизонтальними потоками, тому тип ЛТС називається позиційно-динамічним. Елементиструктури та їх типи. Територіальні одиниці,що оточують каркасні лінії динаміки ландшафту, здебільшого мають форму смуг. Тому Г. І. Швебс і назвав їх ландшафтними смугами. Ландшафтна смуга— це група геотопів, які мають спільне положення в ідносно меж зміни інтенсивності горизонтальних речовинно-енергетичних потоків (розташовані між двома суміжними каркасними лініями динаміки ландшафту). У межах однієї ландшафтноїсмуги горизонтальні потоки односпрямовані і в усіх геотопах мають однаковий градієнт. Оскільки в ландшафтній смузі мають бути однаковими градієнти горизонтальних динамічних потоків, то межі між ними проводяться по каркасних лініях рельєфу. Важливою ознакою ландшафтної смуги є також однорідність її грунтів у протиерозійному відношенні та характером мікрорельєфу. Л андшафтні яруси — одиниці позиційно-динамічної ЛТС, які складаються з групи територіально суміжних, пов'язаних односпрямованими горизонтальними потоками ландшафтних смуг, що мають спільну дозицію щодо гіпсометричних меж зміни провідних факторів ландшафтної динаміки. Ландшафтні яруси відрізняються не тільки висотним положенням, а насамперед набором ландшафтно-екологічних процесів. Ландшафтні яруси пов'язані між собою горизонтальними односпрямованими потоками об'єднують у вищуодиницю позиційно--дииамічної ЛТС— парадинамічний район –сукупність ландшафтних ярусів, пов'язаних горизонтальними потоками які починаються від спільного «центрального місця» —ярусу, що займає панівне висотне положення. Ландшафтні яруси в одному парадинамічному районі, що мають одну макроекспозицію, можна об'єднати в парадинамічний район і. Ці одиниці здебільшого збігаються з частинами (правоюі лівою) басейнів річок, розділених спільним вододілом. 3. Парагенетична ландшафтна територіальна структура. Структуроформуючі відношення. Парагенетична ЛТС формується вздовж ліній концентрації речовинно-енергетичних потоків (наприклад, вздовж річкового русла), ліній розділу контрастних середовищ (берегова лінія) та деяких інших «центральних місць», які визначають напрямок ландшафтогенезу. Такі структури недалеко від «ядра збурення» мають більш чітку парагенетичну диференціацію, яка з віддаленістю від нього послаблюється. Особливе значення у виділенні парагенетичної ЛТС має концентрований водний потік. Його динамічність та енергія, особливі властивості води як природного тіла зумовлюють утворення ландшафтних структур з добре виявленими парагенетичними відношеннями між геотопами. Ці відношення найчіткіше проявляється вздовж ліній потоку і послаблюються з віддаленістю від них. В територіальні одиниці парагенетичної ЛТС об'єднують і одиниці позиційно-динамічної ЛТС — ландшафтні смуги. Підставою для об'єднання є відношення ландшафтних смуг не до ліній розділу напрямку потоків (як при виділенні парадинамічних районів), а до ліній їх концентрації — річкового русла, тальвегу балки тощо. Таким чином, долинно-річкові, яружно-балкові та подібні їм ст руктури являють собою ніби деталі позиційно-динамічних ланд-ш.'іфтних структур, приурочених до гідрографічної або ерозійної мереж. Територіальними одиницями парагенетичної ЛТС є парагенетичні ландшафтні комплекси (ПГЛК). Комплексом нижчого рангу є парагенетична ланка (ПГ-ланка). Вона являє собою сукупність взаємопов'язаних геотопів, об'єднаних за генетичною єдністю та односпрямованістю розвитку в межах заплавно-руслової частини долини (для долинно-річкових ПГЛК) або днищ ерозійних форм (для яружно-балкових ПГЛК). Закономірне поєднання послідовно спряжених ПГ-ланок та ландшафтних смуг, які спираються на русло, формує ПГ-сектор. ПГ-сектор являє собою ділянку долини однотипну за структурою та динамікою насамперед терас та схилів. ПГ-пояс розглядається як територіально цілісний фрагмент долини з однотипним протіканням сучасних фізико-географічних процесів умовах однієї морфоструктури. Основними ознаками виділення ПГ-поясу є тип долини, особливості морфоструктури, тип рельєфу, стадія розвитку долини, знак та інтенсивність неотектонічних рухів, зональний тип рослинно-грунтового покриву. Згідно з типом «центрального місця», відносно якого виділяється парагенетична ЛТС, розрізняються долинно-річкові, яружно-балкові, прибережно-аквальні, лиманно-гирлові ПГЛК.
Тема: V Ландшафтні територіальні структури (хорологічна ландшафтна екологія) 1. Басейнова ландшафтна територіальна структура. 2. Біоцентрично-сітьова ландшафтна структура. 3. Межі між геосистемами. 4. Способи опису ландшафтних територіальних структур.
1. Басейнова ландшафтна територіальна структура. В. В. Докучаєв та О. І. Воєйков пропонували адміністративні межі проводити по басейнах річок. Як екологічно своєрідні та цілісні системи річкові басейни розглядали В.В. Альохін (1921) М. А. Мензбір (1926). У працях Р. Чорлі—Б. Кеннеді (1971), Я. Демека (1974), Л. М. Коритного (1974), К. М. Дьяконова (1975), О. Ю. Ретеюма (1975), Ф. М. Мількова (1981) стверджувалося, що річковий басейн — не тільки гідрологічна, а й географічна система (геосистема) і об'єкт ландшафтних досліджень. Ю. Одум (1975) пропонував вважати річковий басейн за «мінімальну територіальну одиницю» екосистеми. Структуроформуючі відношення. Концентрований поверхневий стік води з розчиненими та завислими в ній речовинами є структуроформуючим для басейнової ЛТС. Концентрація площинного стоку в лінійний можлива за певної мінімальної площі, з якої поверхневі води збираються до лінійної ерозійної форми. Це призводить до формування басейнів — територій, поверхневі води з яких стікають лише до одного водотоку. Структуроформуючими басейнову ЛТС є не всі водотоки, а лише ті, що мають фіксоване в просторі положення, яке, в свою чергу, визначається глибиною врізу ерозійної форми, а саме річки, сухоріччя, балки, лрщини та яри. Територіальні одиниці та їх типи. Територіальними одиницями басейнової ЛТС є басейни, порядок яких визначає чітку ієрархічну організацію структури в цілому. Крім поділу басейну на його частини за критерієм порядку, в будь-якому басейні можна виділити три підсистеми — долинну,схилову та вододільну. Першу складають днища (для неруслових водотоків), русло, заплава та тераси (для руслових); другу — прирічкові схили; у третій виділяють центральну зону (за Р. Хорто-ном — «пояс відсутності ерозії») та бокову зону межиріч. Заплавні геосистеми відзначаються такими рисами: висотна диференціацію (низька, середня, висока заплави); поперечна зональність (прируслова, центральна, притерасова частини); часова контрастність (різкі зміни водного, ландшафтно-геохімічного та інших режимів); значна біологічна продуктивність; активність формо- та видоутворення рослин і тварин. Надзаплавні тераси в басейновій ЛТС відіграють роль своєрідного гальма (бар'єру) у масоенергообміні між вододільною та долинною підсистемами. Інтенсивні горизонтальні потоки на схилах терас при виході на площадку тераси різко гальмуються і вздовж тилового шва тераси активно акумулюється матеріал, виклинюються грунтові води тощо. До русла цей матеріал не доходить. Схилова підсистема відіграє в басейновій ЛТС не меншу роль, ніж її ядро —водотік. Від набору та інтенсивності схилових процесів залежать об'єм, мутність, хімічний склад річкових вод, їх забрудненість. Вони виступають рефугіумами (притулком) для багатьох видів тварин і рослин. Серед вододільно-рівнинних підсистем за специфікою їх зв'язку з річковою долиною виділяються такі типи (Ф. М. Мільков, 1986): слабко диференційовані вододіли приморських рівнин з практично невираженими зв'язками з річковою долиною; пласкі міжріччя низовин з послабленими зв'язками з річковою долиною; хвилясті асиметричні підвищені рівнини з чіткими зв'язками з долиною; горбисті рівнини з накладеними льодовиково-акумулятивними формами. Типологія басейнів з ландшафтно-екологічних позицій За типом водотоку: річкові, сухорічні, балкові, яружні, лощинні басейни За часткою площі вододільно-рівнинної підсистеми вузькоплакорні та широкоплакорні За часткою лісовкритої поверхні виділяють високозалісені (75—100% площі вкрито лісом), відносно заліснені (50—75), середньозалісені (25—50), малозалісені (5—25), практично безлісні (менше 5 %) басейни. Важливою особливістю басейнової ЛТС є ступінь зв'язку її водотоку з вододільно-рівнинною та схиловою підсистемами. Цей зв'язок визначає залежність хімічного складу річкової води, ступінь її забрудненості, мутності, величини стоку від ландшафтної структури та екологічного стану басейну.. Ступінь зв'язку водотоку з басейном визначається наявністю1 та шириною терас, стрімкістю, формою та довжиною схилів, типом зв'язку річкових вод з ґрунтовими, характером сучасної рослинності, особливо наявністю прирічкових водоохоронних смуг. Оскільки зв'язок водотоку з басейном визначається багатьма ландшафтио-єкологічшши факторами, його можна розглядати як комплексну характеристику басейнової ЛТС (табл. 12). 2. Біоцентрично-сітьова ландшафтна структура. Структуроформуючі відношення. Відношення, які формують даний тип ЛТС, пов'язані з вираженими на хоричному рівні територіальними особливостями поведінки, міграції та взаємовідношень популяцій. Просторові зв'язки між біотичними елементами геосистеми, зумовлені такими процесами, як алелопатія, конкуренція за місцеві ресурси, мають малий радіус дії (до кількох метрів) і зумовлюють формування субтопічної структури геотопу (зокрема, поліекоїдів). На хоричному рівні просторові біотичні відношення реалізуються в таких процесах, як перехресне опилення рослин, рознесення спор, насіння, міграції рослин, тварин. Ці відношення, які пов'язують між собою не окремі живі організми, а їх ценопопуляції та геотопи в цілому, і визначають біоцентрнчно-сітьову структуру ландшафту. Важливими елементами цієї структури є біоцентри — ареали, зайняті геотопами з природною рослинністю, які в антропізованому ландшафті мають відігравати функцію збереження генофонду. Ефективно ця функція може виконуватись у тому разі, якщо площа біоцентру забезпечуватиме умови самовідтворення популяцій, виключатиме можливість їх деградації і вимирання видів внаслідок їх замкненого існування. Як видно з натурних спостережень та результатів численних реалізацій моделей вимирання, площа окремого біоцентру, необхідна для розміщення мінімальної життєздатної популяції ссавців, становить від 10—102 км2 (землерийка, миша) до 104—105 км2 (олень, ведмідь). Для популяцій деревних рослин ці значення менші, проте становлять кілька десятків та сотень квадратних кілометрів. Якщо окремі біоцентри сполучити коридорами, вздовж яких можливий обмін видами та особинами, то є надія значно знизити ймовірність вимирання популяцій, підвищити їх генетичну мінливість та здатність до адаптації, зменшити залежність від катастрофічних змін едафотопів окремих біоцентрів. Для цього необхіднавраховувати територіальні закономірності основних етапів розвитку рослин: генеративного розмноження, особливо опилення та запліднення; дисемінації (розповсюдження насіння); ецезису (приживання рослин на новому місці). Територіальні особливості запилення та запліднення. Для забезпечення виживання виду необхідне перехресне запилення. Особливого значення воно набуває для біоцентрів із незначною чисельністю популяцій, для яких бажаний міжбіоцентричний обмін пилком. Тому важливо визначити відстань, на яку може переноситись пилок. За способом цього переносу виділяють анемофільні види (пилок їх переноситься вітром) та зоофільні (комахами). У помірному поясі комахи переносять пилок на відстань 1,5— 2,0 км, а оптимальний радіус — 800 м, причому найкращими запилювачами вважаються бджоли та джмелі. У лісах помірного поясу серед рослин переважає анемофілія. І. Ф. Удра (1988), узагальнивши дані про перенесення пилку вітром, дійшов висновку, що дальність переносу для дерев не перевищує 300—500 м, а для злаків 200—300 м. А взагалі дальність переносу основної маси пилку дерев дорівнює 3—5-кратній висоті дерева'. Територіальні особливості дисемінації. Рослини переміщуються по території за допомогою спор, насіння, плодів, вегетативних органів. Міграції вегетативним розмноженням відбуваються вкрай повільно — до кількох сантиметрів за рік (наприклад, чорниця — з швидкістю 3—5 см/рік). Насінні ж рослини здатні розносити насіння та плоди на значно більші відстані...';;- Основні^.способи розповсюдження насіння: барохорний (опадання насіння під силою тяжіння), анемохорний (перенесення вітром), зоохорний (тваринами), гідрохорний (поверхневими водами). та антропохорний (завдяки діяльності людини). більшістьрослин при розселенні використовує кілька цих способів. При барохорному розселенні насіння та плоди рослин у межах екоїда. Анемохорне - малий розмір і легкість насінини (осика, береза), крилоподібні плівки (клен, ясен), парусні пристрої (граб, липа), за допомогою яких насіння може розноситись вітром на кілька кілометрів (осики — 2 км, берези — 1,5, листяних порід—до 1 км, хвойних 500—600 м). Водночас, важкі насінини граба, липи, клена, ясена, а також жолуді дуба, горішки бука розносяться вітром не більш як на кілька десятків метрів. Зоохорний спосіб (200—300 м) рознесення насіння птахами, ссавці, кабани – дуб. Характерною особливістю зоохорного переносу є його каналізованість — ссавці і багато видів птахів здебільшого не залишають геотопів з природною і близькою до неї рослинністю (лісосмуги, сади) і переносять насіння переважно в їх межах. Гідрохорний спосіб міграції можливий лише для рослин, насінини яких мають спеціальні морфологічні пристосування до плавання— повітряні порожнини, виділення водовідштовхуючих речовин, щільну структуру тощо. Такі особливості є здебільшого в прибережно-водних, болотяних та деяких видів лучних рослин. їх насіння переноситься водним потоком на відстані до кількох кілометрів. Антропохорне розселення рослин — найефективний засіб розповсюдження на значні відстані. До того ж антропохорно вдається організувати розселення рослин у бажаному напрямку, на бажані відстані і, зокрема, штучно з'єднувати ізольовані біоцентрн біокоридорами. Екологічні закономірності ецезису. Про міграцію певного виду можна твердити лише після того, як пройде його ецезис, тобто проростуть насінини кількох особин виду, нормально розвинуться сходи, пройде вергінільна стадія росту аж до початку плодоносіння та появи нового потомства. Успіх ецезису залежить від придатності для виду екологічних умов геотопу, в який занесено насіння, та його здатності витримати конкуренцію з боку видів-аборигенів. Для ецезису будь-якого виду непридатні орні землі, міська та промислова забудови. Тому міграція між біоцентрами, ізольованими на більш ніж 500—1000 м, можлива лише вздовж біокоридо-рів — природних (залужених або залісених долин, балок тощо) чи антропогенних (лісосмуг, алей у місті). У кожному біокоридорі свої едафічні умови, які визначають екологічні групи рослин, щ® можуть розселюватися вздовж нього. Елементи структури та їх типи. Біоцентрично-сітьову ЛТС складають біоцентри, біокоридори та інтерактивні елементи. Біоцентр — це група суміжних геотопів з природною рослинністю, які виконують функції збереження генофонду ландшафту, оптимізуючого впливу на прилеглі геотопи з культурною рослинністю (рілля) або позбавлені її (міська забудова), естетичної привабливості території. В умовах агроландшафту біоцентрами є окремі гаї, ліси, ділянки степів, луків, боліт, а в міському ландшафті — парки, лісопарки, сквери, райони приватної забудови з присадибними садовими та парковими ділянками. Найчастіше в одному біоцентрі є геотопи з різними едафічнимиумовами. Такою характеристикою може бути едафограма біоцентру (див. рис. 24). Це променева діаграма, осі якої відповідають певним екологічним факторам (зволоженню, засоленості тощо), ступінь впливу яких на біоценоз оцінюється в градаціях від 0 (фактор не діє) до 10 (вплив фактора максимально інтенсивний).
-За біогеографічним значенням чеські ландшафтні екологи виділяють біоцентри локального, регіонального, надрегіонального, провінційного та біосферного рівнів. Біокоридор — видовжений ареал, представлений геотопами з природною або близькою до неї рослинністю, вздовж якого відбуваються біотичні міграції між окремими біоцентрами. У агро-ландшафті біокоридорами є залісені або залужені схили та днища лінійних ерозійних форм, лісосмуги, водоохоронні зони річок, самі річкові долини і взагалі будь-які видовжені ареали, що не розорюються, не зазнають надмірного випасу і щорічного косіння. У міському ландшафті функції біокоридорів можуть виконувати алеї, бульвари, вулиці, ступінь озеленення яких дає змогу мігрувати птахам та комахам між міськими біоцентрами. Забезпечення умов міграції видів—основна функція біокоридору. Важливими функціями є також бар'єрна (снігозатримання, зменшення швидкості поверхневого стоку тощо), екотопічна (місце проживання багатьох видів рослин і тварин, особливо птахів лісостепу), оптимізуючого впливу на прилеглі геотопи, естетична. Біокоридори розрізняються: за генезисом (природні, штучні), місцеположенням (рівнинні, схилові, долинні, балкові, літоральні тощо), едафічними умовами (типізуються й характеризуються аналогічно біоцентрам). За шириною Р. Форман (1983) виділяє лінійні біокоридори — настільки вузькі, що практично не впливають на прилеглі угіддя, та смугасті, ширина яких дозволяє сформуватись у його внутрішній частині специфічним екологічним умовам. Інтерактивний елемент — лінійний ареал, зайнятий геотопами з природною або близькою до неї рослинністю; який відгалужується від біоцентру або біокоридору і виконує функцію поширення їх дії на прилеглі агро- або урбоугіддя. Інтерактивний елемент відрізняється від біокоридору тим, що не з'єднує біоцентрів між собою. Позитивний вплив елементів біоцентрично-сітьової ЛТС проявляється у зменшенні швидкості вітру, збереженні роси, збільшенні вологості грунту та повітря, затриманні та більш рівномірному розподілі снігу на полях, збільшенні видового складу та чисельності ворогів сільськогосподарських шкідників. Зона позитивного впливу біотичних елементів, становить» у середньому 300—500 м. Важливою особливістю біоцентрично-сітьової ЛТС є можливість її планомірної трансформації і навіть створення — штучне формування нових біоцентрів, сполученая їх новими біокоридора-ми, введення інтерактивних елементів тощо. 3. Межі між геосистемами. Сама ж концепція континууму та континуального (плавкого) характеру природних меж зародилася в фітоценології (Л. Г. Раменський): дискретні, тобто лінійні, межі в природі є, але вони зустрічаються рідко, з'являються внаслідок катастрофічних змін едафічних факторів і існують недовго, трансформуючись у перехідні смуги — екотони, або клінальні екосистеми. Типи ландшафтних меж. Одна геосистема іншою може змінюватись двома шляхами: 1) стрибкоподібно вздовж певної лінії (лінійна, або дискретна межа); 2) займати деяку перехідну смугу, межі якої можна визначити (ландшафтний екотон). Прикладом меж є вододільні лінії у різкопочленованому ландшафті, тальвеги, екотони (перехідні смуги) на межі луків та лісу тощо. Умовні межі виділяють якдеякі лінії на реальній перехідній смузі, коли її необхідно умовно зобразити у вигляді лінії (наприклад, на картах). Межі, що розділяють між собою дві елементарні геосистеми (геотопи), можна назвати топічними ландшафтними межами, або межами 1-го рангу. Межі між геосистемами вищих рангів (мікро-геохор, ландшафтних смуг тощо) мають ранг, відповідний рангу контактуючих геосистем. Ранг межі пов'язаний з її контрастністю. Загалом, чим вищий ранг межі, тим вона контрастніша. Контрастність межі — можна оцінити за таким показником:
Морфологічна вираженість меж розділяється на морфологічно невиражені (наприклад, між геосистемами, що різняться між собою лише за ґрунтовими ознаками); слабковиражені (між рівнинними та полого-схиловими геосистемами, вододільні лінії на рівнинних); середньо-виражені (між увігнутою та опуклою частинами схилу); явно морфологічно виражені (бровки схилів, уступи та тилові шви терас). Морфологічна вираженість ландшафтних меж залежить від часу відособлення геосистем. Чим молодша територія, тим чіткіші межі (наприклад, у горах альпійської складчастості, у молодому ерозійному рельєфі). За генезисом, тобто головним фактором, що зумовив появу межі, вони є літогенні, морфогенні, педогенні, гідрогенні, фітогенніг зоогенні, антропогенні. Проте переважна більшість їх комплексні (наприклад, морфолітогенні, педофітогенні тощо). За функцією у ландшафтній територіальній структурі межі бувають контактними та бар'єрними (Каганський, 1982; Родоман,. 1982). Вздовж контактних меж відбувається взаємодія двох сусідніх геосистем, взаємопроникнення їх властивостей, перенесення речовинно-енергетичних потоків. Такими є, зокрема, межі ландшафтних смуг на схилі, межі біоцентрів, ПГ-комплексів. Бар'єрна межа перешкоджає або повністю виключає взаємодію сусідніх геосистем. Щодо напрямку горизонтальних динамічних потоків межі поділяються на дивергентні (межі розсіювання) — від них потоки розходяться у різних напрямках (наприклад, вододільні лінії); конвергентні (межі-концентратори), вздовж яких різноспрямовапі потоки зливаються (наприклад, лінії тальвегів); консеквентні — межі, які збігаються з лініями току (наприклад, лінії скапи СХМ лів); градієнтні, вздовж яких помітно змінюється інтенсивність потоку (наприклад, лінії перегинів схилу, межі ландшафтних смуг) поділяються на градієнтні імпульсні (вздовж них інтенсивність потоку зростає) та градієнтні гальмуючі (зменшується). Ландшафтні екотони. Термін «екотон» увів Клементс у 1928 р. Він розумів під ним перехідну смугу між двома досить контрасними екосистемами, через своєрідність якої їх не можна віднести ні до однієї з суміжних екосистем Виникнення ландшафтних екотонів зумовлене поступовим стиранням різких відмінностей (меж) у природі внаслідок все більшого розвитку міжгеосистемних взаємодій. Здебільшого лінійна межа з часом трансформується в перехідну смугу (чітко виражені бровки молодої ерозійної форми з часом руйнуються і стають все більш пологими поверхнями; лінійна межа між лісом і луками поступо*-во згладжується взаємопроникненням лісових та лучних видів у суміжні біотопи і утворення галявин). Подальший розвиток ландшафтного екотону призводить до формування в ньому деяких специфічних рис, не властивих жодній з контактуючих геосистем. З часом він перетворюється на клінальну, або «типову», геосистому з власними межами, можливо, також екотонного характеру. Переважна більшість меж у ландшафті — екотони різної ширини. За периметром та розміром контактуючих геосистем естои-■ські геоекологи Ю. Мандер та Ю. Ягомягі (1982) виділяють мік-роекотони (вони утворюються при контакті окремих парцел і геотопів до 40 м у діаметрі), мезоекотони (контакт ліс — луки, болото— ліс та ін.), макроекотони (виникають на межі великих лісових, болотних масивів, великих водойм тощо). Г. Вальтер (1976) виділяє також зоно-екотони (наприклад, зона лісостепу, субальпійський пояс), проте цими одиницями ландшафтна екологія здебільшого не займається. Найкраще екотонні ефекти проявляються на контакті лісових геосистем з трав'яними (лучними, степовими, ріллею). Екотоп між лісом та степом відіграє бар'єрну і контактну функції. Бар'єрна може реалізуватись трьома способами: 1) екотон як бар'єр-траисформатор (наприклад, атмосферні потоки тепла і водяної пари в напрямку з лісу до степу в межах екотону нагріваються і висушуються, а в зворотному охолоджуються і зволожуються); 2) екотон як бар'єр-перешкода повітряне перенесення насіння, опаду, перенесення снігу хуртелицею, води поверхневого стоку тощо; 3) екотон як бар'єр-відштовхувач повертає горизонтальні потоки, які йдуть до нього від ядер суміжних геосистем (наприклад, зоогенні міграції типово лісових або типово степових видів). Контактна функція ландшафтного екотону може бути реалізована у вигляді: простого контакту, коли горизонтальні потоки без перешкоди і видозміни перетинають екотон; активного контакту, коли в екотоні формуються нові потоки, невластиві ядрам типовості контактуючих геосистем (наприклад, атмосферні потоки на галявині—«бризові лісові вітри»); вторинного контакту, який проявляється в тому, що матеріал, накопичений в екотоні, починає мігрувати за його межі до суміжних геосистем. Виділивши ландшафтний екотон між двома геосистемами, виділяють і їх ядра типовості (екотон відтинає ці ядра). Визначати місцеположення ядер типовості потрібно при дослідженні ландшафтної динаміки, обгрунтування оптимального розташування пунктів моніторингових спостережень та інших тестових ділянок ландшафтно-екологічних досліджень. Відносно цього ядра можна визначити ступінь типовості будь-якої точки геосистеми: П
4. Способи опису ландшафтних територіальних структур. Картографічний спосіб. Ландшафтні карти.
Графічні та матричні моделі. ЛТС будь-якого типу можна зобразити графом, вершинами якого є геосистєми, а ребрами — просторові відношення між ними. Для генетико-морфологічної ЛТС визначальними є відношення генетичної спільності між її геосистемами.. Обчисливши значення коефіцієнта для всіх можливих пар геосистем, отримаємо матрицю їх близькості - граф подібності (рис. 27). У графічних моделях позиційно-динамічної ЛТС ребра між вершинами (ландшафтними смугами або ярусами) проводяться, якщо між суміжними геосистемами проходить горизонтальний потік Набагато інформативнішими можуть бути матричні та графічні моделі позиційно-динамічної ЛТС, якщо є дані про інтенсивність міжгеосистемних зв'язків — швидкість або масу перенесення через ландшафтну межу певної речовини. Проте, користуючись моделлю міграції (наприклад, площинного стоку), для кожної ландшафтної смуги можна розрахувати масу речовини (наприклад грунту), що змивається з одиниці площі за певний час. Маючи такі оцінки, можна орієнтовно визначити і масу речовини Мц, яка переноситься з і-ої до /-ої геосистеми за цей час:
Обчисливши ці значення для всіх пар суміжних ландшафти* •смуг, можна побудувати граф, для кожної вершини якого легко визначити й баланс надходження — виносу речовини (грунту, радіонуклідів тощо). Приклад такого графу показано на рис. 29.
Графи басейнової ЛТС. Для ландшафтно-екологічного аналізу більш зручні графи, ребрами яких є не водотоки, а відношення підпорядкування басейнів, які показуються на графі як його вершини. У графах біоцентрично-сітьової ЛТС вершинами є біоцентри, ребрами — біокоридори та інтерактивні елементи.. Якщо ж емпіричних даних про інтенсивність міграції немає, її орієнтовно можна визначити за гравітаційною моделлю
Кількісні показники. Для генетик о-м орфологічної ЛТС важливими є показники ступеня її однорідності — різноманітності. Доцільно розрізняти хорологічну різноманітність певного регіону, яка визначається кількістю контурів геосистем у ньому, та типологічну різноманітність, як функцію від кількості видів геосистем у регіоні. Складність ЛТС — її характеристика, що поєднуйоцінки хорологічної та типологічної різноманітностей. Контрастність характеризує ступінь несхожості двох сусідніх геосистем або всієї їх сукупності в межах певного регіону. Контрастність можна оцінювати не за всіма ознаками,, що характеризують геосистеми, а за певною їх групою: зволоженням геосистем, їх рельєфом, літологією, засоленням тощо.
. Можна також оцінити показники однорідності — різноманітності по з и ц і й н о-д инамічної ЛТС. Для цього типу ЛТС вони дають певне уявлення про ступінь розвитку сучасних динамічних процесів у регіоні. У загальному випадку чим вища різно-манітність позиційно-динамічної ЛТС, тим інтенсивніші сучасні процеси. Важливою характеристикою парадинамічного району або його ландшафтних ярусів є ступінь зв'язності їх ландшафтних смуг горизонтальними потоками. Кількісно її можна оцінити за допомогою індексу, запропонованого іспанськими екологами Р. Маргалефом та Е. Гутієрес (1983) для оцінки зв'язності трофічної структури екосистеми. Інтерпретація цього індексу до ЛТС має вигляд
Основною функцією біоцентричної сітьової ЛТС є забезпечення біотичної різноманітності та генофонду шляхом міграції видів. Тому важливо кількісно оцінити, на скільки ефективно ця функція може бути виконана. Деякі уявлення про це дають прості показники — кількість біоцентрів, їх розмір, довжина біокоридорів, процент площі, який займають біоцентри та біо-коридори на певній території тощо. Проте більш інформативними «показники, які характеризують ступінь зв'язності графу. Стосовно графу біоцентрично-сітьової ЛТС американські (Р. Формам, М. Годрон) та чеські (М. Козова та ін.) ландшафтні екологи їж-
Тема VI. ГЕОСИСТЕМИ ТА ЇХ СЕРЕДОВИЩЕ (факторіальна ландшафтна екологія) План. 1. Природно-ландшафтні екологічні фактори. 2. Концепція ландшафтно-екологічної ніші. 3. Об¢єм та перекриття ніш. 1. Природно-ландшафтні екологічні фактори. Типологія факторів. Під фактором розуміється деяка ознака, що впливає на характеристики досліджуваної системи. Якщо ця ознака характеризує не саму цю систему, а належить до її зовнішнього середовища, то фактор називається зовнішнім, або екзогенетичним (кількість сонячної радіації, що надходить, кількість та інтенсивність атмосферних опадів, знак та швидкість сучасних тектонічних рухів). Внутрішні, або едногенетичні, фактори співвідносяться з властивостями самої геосистеми, їх речовинний носій знаходиться в межах її вертикальної або територіальної структури і визначає зміни деяких зв'язаних з ним інших елементів геосистеми (засоленість літогеогоризонтів, стрімкість схилу, вміст гумусу в грунті, щільність та біомаса популяцій) Прийняття тієї чи іншої ознаки геосистеми за її внутрішній фактор пов'язане з модульним (моноцентричним, екологічним) підходом до її аналізу, коли певний компонент геосистеми ставиться в центр моделі геосистеми, а інші розглядаються як такі, що визначають його стан та зміни. В екології, а також у ландшафтознавстві досить розповсюджений поділ факторів на абіотичні (кліматичні, орографічні, хімічні тощо) та біотичні, пов'язані з дією на елементи геосистеми * живих організмів. Фактори, що діють на геосистему, доцільніше поділяти на фактори-ресурси та фактори-регулятори. До перших належать тепло, освітленість, кількість опадів, вологи, поживних речовин у грунті тощо. Характер їх впливу визначається кількістю, регулярністю та інтенсивністю надходження до геосистеми (для зовнішніх факторів) та кількістю в самій геосистемі (для внутрішніх). За деякі з цих факторів може йти конкуренція між популяціями і навіть можна вести мову про конкуренцію за ресурс між окремими геокомпонентами (наприклад, за вологу між грунтом та рослинністю), Фактори-регулятори тектонічні рухи, географічне положення геосистеми, рельєф зумовлюють перерозподіл між окремими геосистемами або елементами їх вертикальної структури дії факторів-ресурсів: рельєф визначає неоднакове надходження тепла на схили різних експозицій (при стрімкості схилу 20° ця різниця може сягати 30'%), їх різну освітленість, зволоженість тощо. В результаті на різних елементах рельєфу геотопи мають різну забезпеченість факторами-ресурсами. Тектонічні рухи як зовнішній фактор формування та динаміки геосистем проявляються опосередковано через регуляцію процесів рельєфоутворення, співвідношення виносу та акумуляції матеріалу, глибини залягання рівня ґрунтових вод тощо. Деякі з факторів-ресурсів проявляються і як фактори-регулятори. Так, вміст гумусу діє не тількі як ресурсний фактор для рослин-автотрофів, а й як фактор, що визначає інтенсивність споживання ними елементів мінерального живлення, зокрема добрив. Причому згідно із сучасними уявленнями ця його функція визнається навіть за важливішу, ніж ресурсна. Ландшафтно-екологічна амплітуда. Кожний тип геосистеми може формуватися та існувати лише в деякому діапазоні впливу фактора. Так, на території України геосистеми з чорноземними грунтами зустрічаються лише в діапазоні зволоження від 340 до 600 мм, із сірими лісовими 460—660 мм на рік. За фактором тепла (річного радіаційного балансу) ці діапазони відповідно становлять 41—56 та 40—46 ккал/см2 на рік. Поза цими діапазонами геосистеми даних типів не зустрічаються і заміщуються іншими. В екології діапазон впливу фактора, в якому може існувати певний вид, називається амплітудою виду. Причому розрізняють екологічну амплітуду — діапазон існування виду в умовах міжвидової конкуренції, та фізіологічну (або потенційну)—діапазон, який міг би зайняти вид за відсутності конкуренції за ресурс (X. Еленеберг, 1953). Фізіологічна амплітуда завжди ширша за екологічну. Залежно від впливу зовнішніх факторів виділяють ландшафтно-екологічні амплітуди геосистем. Аналогом фізіологічної амплітуди виду може бути глобальна ландшафтно-екологічна амплітуда геосистеми – той діапазон фактора, в межах якого на всій планеті зустрічається певний тип геосистеми. Поняттям за змістом близьким до екологічної амплітуди виду є регіональна ландшафтно-екологічна амплітуда —діапазон фактора, до якого пристосовані геосистеми цього типу в межах певного регіону (наприклад, Східно-Європейської рівнини). Регіональна амплітуда вужча за глобальну. Так, за даними В. Р. Волобу-єва (1974), геосистеми з чорноземами в масштабі планети в цілому розповсюджені в діапазоні 300—800 мм/рік за фактором зволоження та 15—70 ккал/см2 на рік за фактором' тепла (для території України ці амплітуди становлять 340—600 та 41—56 відповідно). Виявлення для геосистеми ландшафтно-екологічної амплітуди за кожним із факторів дає змогу встановити ті з них, що найбільш ускладнюють її існування в даних умовах, такі фактори можна назвати лімітуючими. їх вплив на геосисте-му полягає в обмеженні як можливостей її формування в даних умовах, так і збереження набутої структури, слабкої стійкості до даного фактора. Ландшафтно-екологічна інтерпретація «закону мінімуму» Ю. Лібіха (1841) та «закону толерантності» В. Шелфорда (1913) зводиться до визначення як лімітуючого того з факторів, значення якого для даної геосистеми знаходяться біля крайніх значень її регіональної ландшафтно-екологічної амплітуди. Різні види геосистем розрізняються за складом лімітуючих їх факторів. Так, в умовах тундри ним є тепло, в хвойних і мішаних лісах — трофність (багатство) грунту. В лісостепу грунти багаті, тому природний потенціал геосистем може лімітуватися фактором зволоження. Геосистеми степів мають два основних лімітуючих фактори — зволоженість та засоленість субстрату. В гірських геосистсмах цю функцію відіграють фактори-регулятори — висота та стрімкість схилу. Поблизу точок мінімальних та максимальних значень ландшафтно-екологічної амплітуди геосистеми її стійкість до зовнішніх впливів здебільшого низька. Значення фактора, при якому геосистема займає найбільшу площу, є оптимальним для неї. 2. Концепція ландшафтно-екологічної ніші. Поняття ніші вважається фундаментальним в екології. (Д. Хатчінсон, 1957, Р. Уіттекер, 1980, Е. Піанка, 1981). Термін «ніша» набув в екології широкого вжитку завдяки працям американського орнітолога Д. Грінелла (1917, 1924), який визначивїї як комплекс факторів, необхідних для існування виду. Дещо в іншому розумінні використовував поняття ніші Ч. Елтон (1927). Він надавав їй функціонального значення і під нішею організму розумів спосіб його життя, зокрема, живлення та відношення до ворогів. Концепцію ніші, що має найбільше число послідовників, запропонував Д. Хатчисон у 1957 р. Ця концепція грунтується на понятті багатомірного простору, кожна вісь якого відповідає іичіному екологічному фактору. Оскільки по кожному з факторів вид характеризується відповідною амплітудою, кінці цих амплітуд визначають той об'єм багатовимірного простору, в якому може існувати вид. Цей об'єм Хатчінсон назвав фундаментальною нішею, і в такому розумінні це поняття може бути задіяне до аналізу геосистем. Щоб визначити фундаментальну ландшафтно-екологічну нішу геосистеми, необхідно встановити деякий набір з п факторів, які визначають її існування та ареал поширення на земній поверхні, і за кожним із них виявити ландшафтно-екологічну амплітуду геосистеми. Графічно фундаментальна ніша зображається у вигляді «-мірного паралелепіпеда, сторони якого відповідають ландшафтно-екологічним амплітудам кожного з факторів. Геосистема може існувати лише в межах її фундаментальної ніші. Частина, тобто об'єм фундаментальної ніші, у межах якого геосистема може існувати за будь-якої комбінації факторів, називається реалізованою ландшафтно-екологічною нішею. Практична реалізація концепції ніші як об'єму в багатовимірному просторі факторів пов'язана з побудовою самого цього простору, тобто у визначенні факторів, Місце, яке займає геосистема в ландшафтно-екологічній ніші (на графіку йому відповідає точка об'єму), свідчить про ступінь
її відповідності умовам зовнішнього середовища — біля меж ніші геосистема знаходиться в екстремальних умовах і її стійкість незначна;ближче до центру ніші формуються оптимальні умови для геосистем даного виду. В екології за оцінку беруть продуктивність виду в даних умовах, народжуваність, ефективність фотосинтезу тощо. По відношенню до геоенстем такою оцінкою може бути частка площі, яка припадає на ту частину загального ареалу геосистеми, яка знаходиться в умовах певної комбінації дії факторів, за якими будується ніша. 3. Об¢єм та перекриття ніш. Різні типи геосистем розрізняються не тільки за положенням ніш у просторі факторів, а й за їх величиною (об'ємами). Геосистеми, що мають великі ніші, можуть існувати в широкому діапазоні дії зовнішніх факторів. Геосистеми ж з малими нішами дуже вибагливі до умов зовнішнього середовища і загалом нестійкі при варіації цих умов. Об'єм ландшафтно-екологічних ніш слід враховувати при визначенні природоохоронних пріоритетів— геосистеми з вузькими нішами потребують особливої уваги та природоохоронного статусу, оскільки такі геосистеми часто є реліктовими або екзотичними для певного регіону, їх стійкість здебільшого невисока. Кількісно об'єм ніші можна оцінити за виразом Vs=П( ΔXi /Xm) де Vs— об'єм ландшафтно-екологічної ніші; ΔXi — величина ландшафтно-екологічної амплітуди геосистеми за 1-м фактором; Xm— центральне значення амплітуди; п — число факторів, за якими побудована ніша (осей її простору); П — знак добутку. Ландшафтно-екологічні ніші окремих геосистем перекриваються. Це свідчить про те, що в одних умовах зовнішнього середовища може формуватися та існувати кілька різних видів геосистем. Геосистеми, ніші яких перекриваються за всіма факторами, близькі за структурою та динамічними тенденціями. Тому оцінка ступеня перекриття ніш є показовою характеристикою подібності геосистем, зумовленою факторами зовнішнього середовища. Перекриття ніш геосистем свідчить також про можливість їх боротьби за простір — витіснення геосистем менш пристосованих до даних умов зовнішнього середовища більш пристосованими. Активність та напрям цієї конкуренції в певних умовах дії зовнішніх факторів (тобто в певній точці простору ніш) визначаються різницею між показниками пристосованості р конкуруючих у даних умовах геосистем. Якщо ландшафтно-екологічні ніші окремих геосистем перекриваються, то точки їх оптимумів індивідуальні для кожного типу геосистеми. Співставивши ландшафтно-екологічні ніші основних зональних типів геосистем України в двовимірному просторі факторів ми виявили, що точки їх оптимумів лежать на прямій, яка також проходить вздовж гребенів ніш усіх геосистем. Цю пряму можна назвати віссю л
|

 структури фітоценозу слід додати четвертий — мікроценотичний. Мікроценози (синонім — мікроугруповання), за П. Д. Ярошенком (1958, 1961), В. І. Василевичем (1973, 1983) та іншими фітоценологами, являють собою гомогенні елементарні частини з яких складається фітоценоз. Контури мікроценозів здебільшого пов'язані з певними внутрішньотопічними відмінностями в грунтах, мікрорельєфі, умовах зволоження. М. В. Диліс (1964), розглядаючи мікроценози з відповідними їм грунтами, мікрокліматом, тваринним населенням тощо, називає їх парцелами і вважає за елементарні частини біогеоценозу (одиниці близької до геотопу). М. В. Диліс вважає важливою ознакою парцел їх відособленість на всю вертикальну товщу біогеоценозу. З цим погодитися важко, бо, наприклад, нижні горизонти грунту та верхні верстви геологічних порід у межах геотопу від варіації рослинності здебільшого не залежать.
структури фітоценозу слід додати четвертий — мікроценотичний. Мікроценози (синонім — мікроугруповання), за П. Д. Ярошенком (1958, 1961), В. І. Василевичем (1973, 1983) та іншими фітоценологами, являють собою гомогенні елементарні частини з яких складається фітоценоз. Контури мікроценозів здебільшого пов'язані з певними внутрішньотопічними відмінностями в грунтах, мікрорельєфі, умовах зволоження. М. В. Диліс (1964), розглядаючи мікроценози з відповідними їм грунтами, мікрокліматом, тваринним населенням тощо, називає їх парцелами і вважає за елементарні частини біогеоценозу (одиниці близької до геотопу). М. В. Диліс вважає важливою ознакою парцел їх відособленість на всю вертикальну товщу біогеоценозу. З цим погодитися важко, бо, наприклад, нижні горизонти грунту та верхні верстви геологічних порід у межах геотопу від варіації рослинності здебільшого не залежать.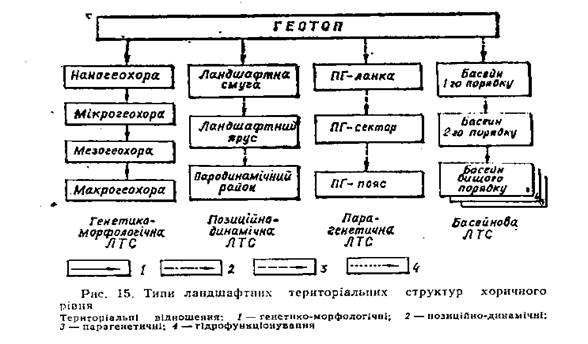



 'За площею виділяються такі біоцентри: карликові (в агроланд-шафті 0,2—0,5 км2; в міському ландшафті — 0,05—0,1 км2); малі (відповідно 0,5—1 і 0,1—0,3), середні (1—3 і 0,3— 1), відносно великі (3—10 і 1—3) та великі (^10 і ^3). За сучасним станом біоцентри поділяються на критичного, пригніченого та нормального розвитку. Критеріями визначення цих типів є процент рослин, уражених шкідниками, з морфологічними ознаками пригніченості, викликаної природними та антропічними фа'кторами, розвиток підросту, частка молодих особин у фітоценозі тощо.
'За площею виділяються такі біоцентри: карликові (в агроланд-шафті 0,2—0,5 км2; в міському ландшафті — 0,05—0,1 км2); малі (відповідно 0,5—1 і 0,1—0,3), середні (1—3 і 0,3— 1), відносно великі (3—10 і 1—3) та великі (^10 і ^3). За сучасним станом біоцентри поділяються на критичного, пригніченого та нормального розвитку. Критеріями визначення цих типів є процент рослин, уражених шкідниками, з морфологічними ознаками пригніченості, викликаної природними та антропічними фа'кторами, розвиток підросту, частка молодих особин у фітоценозі тощо.










 що зумовлюють можливість існування та ареал, що займає геосистема певного типу. Факторами, відносно яких доцільно будувати нішу геосистем, є річна кількість опадів, радіаційний баланс, а в гірських геосистемах — абсолютна висота та макроекспозиція. Для ландшафтно-екологічної ніші геотопів, наногеохор та ландшафтних смуг важливими факторами є крутизна поверхні, довжина схилу, його експозиція, мінералізація та глибина залягання ґрунтових вод. Радіаційний баланс та річна сума опадів у геосистемах цих рангів виступають як константи.
що зумовлюють можливість існування та ареал, що займає геосистема певного типу. Факторами, відносно яких доцільно будувати нішу геосистем, є річна кількість опадів, радіаційний баланс, а в гірських геосистемах — абсолютна висота та макроекспозиція. Для ландшафтно-екологічної ніші геотопів, наногеохор та ландшафтних смуг важливими факторами є крутизна поверхні, довжина схилу, його експозиція, мінералізація та глибина залягання ґрунтових вод. Радіаційний баланс та річна сума опадів у геосистемах цих рангів виступають як константи.



