Динамика популяций
Репродуктивный потенциал и рост популяции. Динамика популяции – это процессы изменений её основных биологических показателей во времени. Особенно большое значение имеет изменение её численности. Любая популяция теоретически способна к неограниченному росту численности, если её не лимитируют факторы среды. В таком случае скорость роста численности популяции будет зависеть только от величины репродуктивного (биотического) потенциала, свойственного виду. Репродуктивный потенциал отражает максимальное число потомков от одной пары (или одной особи) за единицу времени (сутки, год или за весь жизненный цикл). Величина репродуктивного потенциала неодинакова у разных видов и зависит от размеров особей, продолжительности их жизни, уровня эволюционного развития и смертности в популяциях. Корреляция видового уровня плодовитости и средней нормы гибели установлена для различных групп живых организмов. Растения, грибы производят огромное количество спор, так как большинство из них погибает, не попав в благоприятные для развития условия. Рыбы с пелагической икрой, доступной для поедания многими животными, откладывают её в огромном количестве, тогда как виды, укрывающие каким-либо способом свою икру, имеют более низкую плодовитость. Плодовитость сильно зависит от степени развития заботы о потомстве или обеспеченности яиц питательными веществами. Например, акулы, яйца у которых крупные, с большим запасом питательных веществ и имеют плотную защитную оболочку, откладывают несколько яиц за сезон. Зелёная жаба, не заботящаяся о потомстве, производит за сезон 8–12 тысяч икринок, тогда как жаба-повитуха, наматывающая икру на задние лапы, – всего 150 штук. Таким образом, появление в эволюции каких-либо форм заботы о потомстве чётко коррелирует со снижением видовой нормы плодовитости. У таких видов снижается смертность в раннем возрасте и сохранение высокого репродуктивного потенциала биологически не выгодно. Обратная зависимость наблюдается между плодовитостью и средней продолжительностью жизни, свойственной данному виду. Долгоживущие виды отличаются меньшей плодовитостью. В реальной обстановке конкретные величины как плодовитости так и смертности постоянно колеблются, соответственно этому меняются и уровень численности популяции, и тенденции её изменения во времени. Если бы всё потомство выживало, то численность любой популяции увеличивалась бы в геометрической прогрессии. Такой рост выражается экспоненциальной кривой (рис. 10).
В природе биотический потенциал популяции никогда не реализуется полностью. Это может наблюдаться в отдельных случаях в течение короткого периода времени, когда, например, быстро размножающиеся организмы (бактерии, грибы, насекомые) осваивают какой-либо субстрат или среду, где нет конкурентов. Естественный рост популяции никогда не реализуется в форме экспоненциальной модели, так как он ограничен комплексом факторов внешней среды (наличием ресурсов, конкурентов и др.) и реально складывается как результат соотношений меняющихся значений рождаемости и смертности. Наиболее близко естественный рост численности популяций отражает логистическая модель роста, в которой изменение численности во времени выражается S-образной кривой (рис.11).
В соответствии с логистической моделью рост численности популяции некоторое время идёт медленно. Затем кривая численности резко возрастает и наконец выходит на уровень, определяемый ёмкостью среды. Этот конечный уровень отражает уравновешенность процессов рождаемости и смертности в соответствии с наличием пищевых и иных ресурсов среды.
Колебания численности популяции. Рассмотренные выше модели роста численности популяции представляют собой «идеальные» схемы, основанные на видовых особенностях механизма самоподдержания популяции. Сложность условий, в которых функционируют естественные популяции, приводит к тому, что их численность после достижения верхнего уровня роста подвержена изменениям. Такие колебания могут возникать либо в результате изменения физической среды, вследствие чего повышается или снижается верхний предел численности, либо в результате внутрипопуляционных и межпопуляционных взаимодействий. После того как верхний предел численности популяции достигнут, она может некоторое время оставаться на этом уровне (рис. 12, кривая 1) или сразу резко упасть (рис. 12, кривая 2). Возможна и такая ситуация, при которой численность популяции «перескакивает» через уровень (кривые 3, 4). В этом случае кормовые и другие ресурсы, необходимые для жизни популяции, накоплены ещё до начала её роста, а механизмы регуляции численности ещё не действуют. Например, при вселении животных туда, где их раньше не было.
Рис. 12. Различные типы (1–4) колебания численности популяции
Колебания численности популяции могут быть непериодическими (вызванными воздействием каких-либо факторов) и периодическими (циклическими, повторяющимися через определённый период времени). Непериодические колебания численности носят непредвиденный характер. Циклические изменения численности с периодом в несколько лет известны для популяций некоторых арктических млекопитающих и птиц. Например, у зайца-беляка и рыси период колебаний численности равен примерно 9–10 лет (рис. 13).
Рис. 13. Периодические колебания популяций зайца-беляка (1) и рыси (2), установленные по числу шкурок, заготовленных Компанией Гудзонова пролива (из Р. Дажо, 1975) Более короткие, 3- и 4-годичные, циклы характерны для многих обитателей тундры: песца, полярной совы, лемминга, полёвок, лисиц. Подобные колебания численности известны также для многих насекомых (бабочек, саранчи и др.)
Рис. 14. Типы динамики населения по С.А. Северцову: I – стабильный, II – лабильный, III – эфемерный
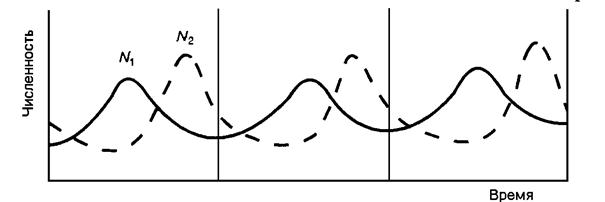
Стабильный тип характеризуется малой амплитудой и длительным периодом колебаний численности (10–29 лет). Он свойствен крупным животным с большой продолжительностью жизни, поздним наступлением половой зрелости и низкой плодовитостью. Это соответствует низкой норме естественной смертности, так как имеются эффективные механизмы адаптации к неблагоприятным факторам. Примером могут служить копытные млекопитающие, китообразные, приматы, орлы и др. Лабильный тип динамики отличается закономерными колебаниями численности с периодами от 5 до 11 лет и более значительной амплитудой (численность меняется в десятки раз). Характерен для животных с продолжительностью жизни 10–15 лет, ранним половым созреванием и более высокой плодовитостью (зайцеобразные, крупные грызуны, многие птицы, рыбы и др.). Эфемерный тип динамики отличается резко неустойчивой численностью: она может возрастать в сотни раз и очень быстро падать. Перепады от максимума до минимума происходят очень быстро, иногда в течение одного сезона. Длина цикла составляет до 4–5 лет, в течение которых «пик» численности занимает не более одного года. Такой тип динамики характерен для видов с большой плодовитостью, высокой нормой гибели и с продолжительностью жизни не более 3 лет (мелкие грызуны, насекомые с коротким циклом развития). Рассмотренные типы динамики показывают связь этих процессов с биологическими особенностями видов и их популяций, отражают разные жизненные стратегии. Эта мысль лежит в основе концепции экологических стратегий, разработанной американскими экологами Э. Мак-Артуром и Э. Уилсоном (1967). Суть этой концепции сводится к тому, что успешное выживание и воспроизводство вида возможно либо путём совершенствования адаптированности организмов и их конкурентоспособности, либо путём интенсификации размножения, что компенсирует повышенную гибель особей и в критических ситуациях позволяет быстро восстановить численность. Первый путь назван «К-стратегией» – это крупные формы с большой продолжительностью жизни, численность их лимитируется преимущественно внешними факторами. К-стратегия означает «отбор на качество», повышение адаптивности и устойчивости; «r-стратегия» – «отбор на количество» через компенсацию больших потерь высокой плодовитостью. Виды r-стратегии легко осваивают местообитания с нестабильными условиями, благодаря высокой плодовитости быстро восстанавливают любые потери в популяции. Аналогичные жизненные стратегии свойственны и растениям. Английский учёный Д. Грайм выделил среди них такие стратегии: конкурентная – достигающая высокой плотности в оптимальных местообитаниях; стресстолерантная – устойчивые к неблагоприятным факторам, но малопродуктивные виды, заселяющие менее благоприятные места; рудеральные – отличаются высоким репродуктивным потенциалом и быстрым ростом, заселяют антропогенные места. Как видно из изложенного, в экологических стратегиях растений и животных обнаруживается большое сходство, что говорит об общности наиболее фундаментальных экологических основ динамики популяций в разных группах живых организмов. Факторы динамики численности. Факторы, ответственные за изменение численности популяций в современной экологии, принято делить на две группы: 1) факторы, не зависящие от плотности популяции; 2) факторы, зависящие от плотности популяции. К первой группе относятся абиотические факторы среды. Они действуют на уровне организма, действие их одностороннее, т.е., организмы не могут оказать на них обратное влияние. Их называют модифицирующими факторами, так как они вызывают изменения численности популяций, но не могут её регулировать. Эффект воздействия абиотических факторов на уровень численности реализуется через изменение рождаемости и смертности. Благоприятные условия могут послужить причиной массовой вспышки размножения вида; а в случае стадных саранчовых неблагоприятные климатические погодные условия, наоборот, снижают численность популяции иногда до полного её исчезновения. Например, в малоснежные холодные зимы в лесной зоне из-за промерзания почвы в массе гибнут кроты, полёвки, мыши и др. Заморозки в начале лета могут повлиять на выживаемость многих организмов. Факторы, зависящие от плотности популяции, не просто изменяют её численность, а сглаживают колебания, приводя после отклонения от оптимума к прежнему уровню. Это происходит потому, что эффект их воздействия тем сильнее, чем выше плотность популяции. Их называют регулирующими факторами, так как они действуют по принципу обратной связи. В качестве регулирующих факторов выступают межвидовые и внутривидовые отношения организмов: возбудители болезней, конкуренция, хищничество, паразитизм, собирательство и др. Одна из важных форм взаимоотношений организмов, влияющая на периодические изменения численности популяций, – отношение потребителя и его пищи. Примером такого рода взаимоотношений могут служить «лемминговые циклы», сопровождающиеся катастрофическими нарушениями тундровой растительности, массовыми миграциями и гибелью животных. Но циклические изменения их численности связаны не только с нехваткой пищи и её последующим восстановлением. Выяснилось, что в период пика численности наблюдаются увеличение агрессии особей, возникновение стресса, что ведёт к снижению рождаемости через механизмы популяционного гомеостаза. Наиболее изучены циклические изменения численности популяций хищников и их жертв. В лабораторных условиях при совместном содержании хищника и его жертвы наблюдаются периодические изменения обоих видов, имеющие довольно закономерный характер (рис. 15).
Рис. 15. Взаимосвязанные колебания численности в системе хищник – жертва по модели Лотки – Вольтерра: N1 – жертва, N2 – хищник
Прежде всего это относится к специализированным хищникам, которые не могут переключиться на другие виды пищи при снижении численности их основной жертвы. Увеличение численности жертв идёт до тех пор, пока темп их выедания нарастающим количеством хищников не превысит скорость их размножения. После этого дальнейшая деятельность хищников подрывает кормовую базу, что ведёт к снижению их собственной численности. Однако в природных условиях такие закономерные циклические изменения численности в системе хищник – жертва выявить трудно, так как реальная динамика численности идёт под влиянием многих факторов. Иногда такие колебания обнаруживаются в бедных сообществах, где межвидовые связи не столь разнообразны: в лесах с господством одной древесной породы, в тундре, арктических пустынях и др. Примерно по такой же схеме происходит динамика численности популяций при взаимодействии паразитов и возбудителей болезней с их хозяевами. При повышении плотности популяций возрастает вероятность заражения возбудителями болезней. В результате вспышки заболевания происходит вымирание животных, что ведёт к снижению численности возбудителя, который в выживших животных интенсивно размножается, увеличивая свою численность, и вновь вызывает вспышку заболевания. Однако в природе такие закономерные колебания численности в рассмотренных системах паразит – хозяин и возбудитель болезни – хозяин часто осуществляются на протяжении короткого промежутка времени, после чего следует отклонение от классической схемы. В связи с этим ряд учёных (Н.Н. Наумов, А.А. Максимов) полагают, что биоценотические взаимоотношения организмов лишь сдерживают чрезмерный рост численности популяций. Процесс динамики численности популяций очень сложный, в нём невозможно выделить один ведущий фактор или даже небольшое их число. Роль отдельных факторов, определяющих общий уровень численности и тенденции к её изменению, меняется в зависимости от условий местности и состояния популяции в разные сезоны и годы.
|
 Рис. 10. Теоретическая кривая роста популяции
Рис. 10. Теоретическая кривая роста популяции Рис. 11. Логистическая кривая роста популяций
Рис. 11. Логистическая кривая роста популяций 1 2 3 4
1 2 3 4
 Характер закономерных периодических изменений численности популяций видоспецифичен и связан с особенностями биологии вида, его физиологии и места в естественных экосистемах. С.А. Северцов, ещё в 40-х годах прошлого века проанализировавший многолетний ход динамики численности популяций у многих видов млекопитающих и птиц, установил три фундаментальных типа: 1) стабильный, 2) лабильный, 3) эфемерный (рис. 14).
Характер закономерных периодических изменений численности популяций видоспецифичен и связан с особенностями биологии вида, его физиологии и места в естественных экосистемах. С.А. Северцов, ещё в 40-х годах прошлого века проанализировавший многолетний ход динамики численности популяций у многих видов млекопитающих и птиц, установил три фундаментальных типа: 1) стабильный, 2) лабильный, 3) эфемерный (рис. 14).