Экологические стратегии популяций
Экологические стратегии популяций – это общая характеристика роста и размножения в пределах данной популяции. В 1938 году русский ботаник Леонтий Григорьевич Раменский выделил три основных типа стратегий выживания среди растений: - виоленты – подавляют всех конкурентов. К ним относятся деревья, тростники, сфагновые мхи, доминирующие на заболоченных территориях; -патиенты – виды, способные выжить в неблагоприятных условиях («тенелюбивые», «сухолюбивые», «солелюбивые» и т.п.); - эксплеренты – виды, способные быстро появляться там, где нарушены коренные сообщества, – на вырубках и гарях (кипрей, осины, березы), на отмелях и т.д. На основании скорости роста популяций, рождаемости, смертности и прочих характеристик выделяют несколько экологических стратегий популяций: а) r-стратеги (r-виды, r-популяции) – популяции из быстро размножающихся, но менее конкурентоспособных особей. Имеют J-образную (экспоненциальную) кривую роста численности. Такие популяции быстро расселяются, но они малоустойчивы и быстро погибают. К ним относятся бактерии, тли, однолетние растения и др. б) к-стратеги (к-виды, к-популяции) – популяции из медленно размножающихся, но более конкурентоспособных особей. Имеют S-образную кривую роста численности. Такие популяции населяют стабильные местообитания. К ним относятся птицы, млекопитающие, деревья и др.
35. Динамика роста численности популяции. “Кривые“ роста популяции (графическое изображение) и их математические формулы. Скорость роста популяции – это изменение численности популяции в единицу времени. Она может быть либо положительной, либо нулевой, либо отрицательной и зависит от показателей рождаемости, смертности и миграции особей. Различают абсолютную и удельную скорость роста популяции: а) абсолютная (общая) скорость роста – выражается изменением численности популяции за промежуток времени; б) удельная скорость роста – отношение скорости роста к исходной численности. Скорость роста может быть выражена в виде кривой роста популяции (рисунок).
Рисунок – Кривые роста популяции
Существует две основные модели роста популяции: - J-образная кривая – отражает неограниченный экспоненциальный рост численности популяции, не зависящий от плотности популяции. Подобный рост популяций иногда наблюдается в природе: «цветение» воды в результате бурного развития фитопланктона, вспышка массового размножения некоторых вредителей, рост бактерий в свежей культуре. Однако это происходит непродолжительное время, так как после превышения емкости среды неизбежно произойдет резкое снижение численности. - S-образная (сигмоидная, логистическая) кривая – отражает логистический тип роста в реальных экологических условиях, зависящего от плотности популяции, при котором скорость роста популяции снижается по мере роста численности (плотности). Сначала рост популяции невелик, но затем он нарастает, но через некоторое время замедляется и выходит на плато (рисунок).
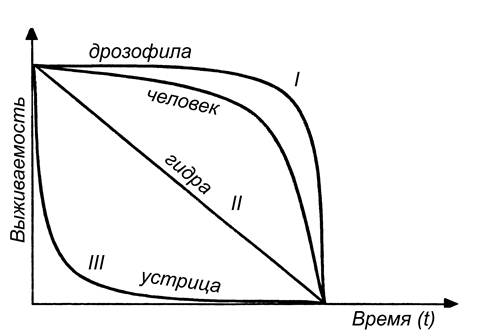
36. Продолжительность жизни и выживаемость. Кривые выживания. Существует 3 типа смертности, которым соответствуют определенные кривые выживаемости (рисунок): Кривая I типа свойственна организмам, смертность которых на протяжении всей жизни незначительна, но возрастает в ее конце. Идеальная кривая для выживаемости популяции, в которой лишь старение служит главным фактором, влияющим на смертность (например, популяции лабораторных животных в идеальных условиях, люди в развитых странах, некоторые крупные млекопитающие) Это так называемая «кривая дрозофилы». Кривая II типа характерна для видов, у которых смертность остается примерно постоянной в течение всей жизни. Это могут быть растущие популяции в оптимальных условиях среды (например, популяции высокоорганизованных птиц, млекопитающих). Это «кривая гидры». Кривая III типа отражает массовую гибель особей в начальный период жизни, причем смертность постоянна в течение всей жизни организмов. Главный фактор, определяющий смертность, – случай («популяция стеклянных стаканов в кафетерии»). Такая кривая характерна для большинства популяций растений и животных (например, многие рыбы, беспозвоночные, растения и другие организмы, не заботящиеся о потомстве, и выживающие за счет огромного количества икринок, личинок, семян и т.п.). Это «кривая устрицы».
Рисунок – Кривые выживаемости
Изучение смертности имеет важное значение для определения степени уязвимости популяций вредителей народного хозяйства.
37. Экологическая стратегия выживания. Основные признаки К- и r-стратегов. Типы стратегий по Л.Г. Раменскому. Экологические стратегии популяций – это общая характеристика роста и размножения в пределах данной популяции. В 1938 году русский ботаник Леонтий Григорьевич Раменский выделил три основных типа стратегий выживания среди растений: - виоленты – подавляют всех конкурентов. К ним относятся деревья, тростники, сфагновые мхи, доминирующие на заболоченных территориях; -патиенты – виды, способные выжить в неблагоприятных условиях («тенелюбивые», «сухолюбивые», «солелюбивые» и т.п.); - эксплеренты – виды, способные быстро появляться там, где нарушены коренные сообщества, – на вырубках и гарях (кипрей, осины, березы), на отмелях и т.д. На основании скорости роста популяций, рождаемости, смертности и прочих характеристик выделяют несколько экологических стратегий популяций: а) r-стратеги (r-виды, r-популяции) – популяции из быстро размножающихся, но менее конкурентоспособных особей. Имеют J-образную (экспоненциальную) кривую роста численности. Такие популяции быстро расселяются, но они малоустойчивы и быстро погибают. К ним относятся бактерии, тли, однолетние растения и др. б) к-стратеги (к-виды, к-популяции) – популяции из медленно размножающихся, но более конкурентоспособных особей. Имеют S-образную кривую роста численности. Такие популяции населяют стабильные местообитания. К ним относятся птицы, млекопитающие, деревья и др.
38. Факторы динамики численности. Факторы, вызывающие изменение численности, разнообразны. Их подразделяют на две группы: не зависимые и зависимые от плотности популяции. К не зависимым от плотности популяции относят преимущественно абиотические факторы. Они действуют на популяцию при любой ее численности. Например, особо суровые зимы вызывают гибель зимующих особей капустной белянки вне зависимости от того, большое или малое количество особей составляет эту популяцию в данный зимний период. Или наоборот, благоприятные условия зимовки могут способствовать повышению численности особей как в малочисленных популяциях, так и в больших. Следовательно, разнообразные абиотические факторы среды могут вызвать значительные колебания численности популяции. К зависимым от плотности популяции принадлежат биотические факторы — естественные враги (хищники, паразиты, возбудители болезней) и пищевые ресурсы. Их количество изменяется вместе с изменением численности популяции. Установлено, что как только плотность популяции того или иного вида хищников увеличивается, численность популяции его основной жертвы начинает снижаться. Такой же эффект на популяцию хозяина оказывают и паразиты. Как правило, чем выше плотность популяции, тем сильнее влияние этих факторов. Без них численность популяции могла бы неограниченно увеличиваться, что привело бы к полному уничтожению источников корма. Таким образом, особенность действия факторов^ зависящих от плотности, заключается в сглаживании резких колебаний численности, благодаря чему численность популяции поддерживается на определенном оптимальном уровне. Одним из механизмов регулирования численности является плодовитость. Она снижается при уменьшении пищевой обеспеченности, которое наблюдается при увеличении численности популяции. Снижение плодовитости особей приводит к понижению рождаемости, а следовательно, к замедлению темпов роста популяции (рис. 1.7). Важную роль в регуляции плотности популяции играют поведенческие факторы, в частности территориальность. Наличие у особей определенного вида своего индивидуального участка, который обозначается разными способами (мочой, секретом пахучих желез, царапинами на деревьях, звуками и др.), ограничивает рост численности их популяций, так как особи, не имеющие собственного участка, не участвуют в размножении. Таким образом, популяции подвержены воздействию комплекса абиотических и биотических факторов, которые приводят в действие механизмы регуляции их численности. Поэтому в не нарушенных деятельностью человека природных сообществах редко происходит неудержимый рост численности, исчерпание ресурсов и гибель популяций. Численность естественных популяций не остается постоянной, так ках изменчивы условия окружающей их среды. Диапазон изменчивости численности различен у разных видов. Он обусловлен степенью изменчивости условий среды обитания, а также биологическими особенностями конкретного вида. Выделяют три наиболее часто встречаемых типа динамики численности: сезонный, многолетний периодический и устойчивый. Регуляция численности в природных условиях осуществляется не зависимыми и зависимыми от плотности популяции факторами.
39. Концепция саморегуляции, стресс-реакция. Множественность механизмов популяционного гомеостаза. Сколь бы ни велики были различия между стохастизмом и регуляционизмом, сторонники этих подходов сходились на том, что ведущая роль в ограничении роста численности популяций принадлежит факторам внешней среды, например таким, как нехватка пищи или неблагоприятные погодные условия. Однако в начале 60-х гг. была предложена завоевавшая вскоре большую популярность концепция саморегуляции популяций, согласно которой в процессе роста плотности популяции изменяется не только и не столько качество среды, в которой существует эта популяция, сколько качество самих составляющих ее особей. Это изменение свойств особей, направленное на то, чтобы затормозить дальнейший рост популяции, выражается в конечном счете - в снижении плодовитости, - удлинении сроков полового созревания, - возрастании смертности и - миграционной активности. Как подчеркнул в свое время один из авторов этой концепции английский эколог Д. Читти (Chitty), любая популяция способна в принципе регулировать свою численность так, чтобы не подрывались возобновляемые ресурсы местообитания и не требовалось вмешательства каких-либо внешних факторов, например хищников или неблагоприятной погоды. Согласно концепции саморегуляции изменение качества особей, сказывающееся на росте численности, может быть как фенотипическим, так и генотипическим. В последнем случае оно проявляется чаще всего как сдвиг в количественном соотношении разных генотипов В рамках подобной концепции теряет всякий смысл традиционное деление факторов на зависимые и не зависимые от плотности. Основанием для выдвижения гипотезы саморегуляции послужили в первую очередь результаты наблюдений за лабораторными популяциями мышей и других грызунов. Выяснилось, что при возрастании плотности популяции, а точнее, частоты контактов между особями (понятно, что частота контакте зависит не только от абсолютной плотности зверьков, но и от того, как устроено их местообитание, в частности, насколько богато оно всевозможными укрытиями) у грызунов возникает состояние стресса, которое характеризуется рядом признаков, в том числе резким повышением активности надпочечников. Гормональные сдвиги, происходящие в организме под влиянием нервного возбуждения, тормозят деятельность половых желез, что в конечном счете приводит к более позднему половому созреванию, снижении плодовитости, а иногда даже к полному прекращению размножения и резорбции зародышей. Кроме того резко возрастает и смертность как непосредственный результат стрессового состояния, а в природных условиях и как результат резко усилившейся миграции животных в новые местообитаниях, где больше риск гибели от самых разнообразных причин. Среди механизмов, обеспечивающих саморегуляцию популяций, очень важное место принадлежит тем, которые непосредственно связаны с особенностями поведения отдельных особей. Поведение может сказываться на величине рождаемости и смертности через физиологические сдвиги в организме (как, например в случае описанной выше стресс-реакции), а может через изменение пространственного распределения особей. Так, если животные защищают определенную территорию от вторжения чужаков, то при возрастании плотности популяции все большее число особей не могут удержать собственную территорию (или вытеснить ранее обосновавшихся конкурентов) и соответственно должны мигрировать в менее благоприятные места, где они чаще гибнут от хищников, нехватки пищи или воздействия абиотических факторов. Помимо поведенческих механизмов в ограничении роста численности важная роль может принадлежать сопряженным с плотностью изменениям генетического состава популяции. Очевидно, такой генетический механизм подразумевает наличие в популяции по крайней мере двух разных генотипов, один из которых имеет преимущество в условиях высокой плотности, а другой - низкой. Например, канадский эколог Ч. Кребс вместе со своими сотрудниками показал (Krebs), что у пенсильванской полевки (Microtus pennsylvanicus) на пиках численности и в периоды депрессий между пиками доминируют разные генотипы: особи одного генотипа быстро размножаются, но плохо выживают при большой скученности (в период подъема численности они, как правило, мигрируют из основного местообитания), а особи другого — лучше переносят повышенную скученность, но характеризуются меньшей плодовитостью. Генетический механизм регуляции численности скорее всего действует в совокупности с каким-нибудь другим, например с прессом хищников. Рассмотрим приведенный Д. Пайментлом (Pitiientel) гипотетический пример такой взаимосвязи. Пусть какое-нибудь растение, поедаемое травоядными животными, имеет ген, контролирующий его выживаемость и одновременно пригодность в качестве пищи для травоядных. Пусть данный ген встречается в форме двух аллелей А и а, причем гомозиготы АА характеризуются высокой выживаемостью и одновременно съедобностью для травоядных, гомозиготы аа - низкой выживаемостью и практической несъедобностью для травоядных, а гетерозиготы Аа отличаются промежуточными свойствами. В итоге соотношение рассматриваемых свойств у трех возможных генотипов нашего гипотетического растения следующее:
Если предположить, что содержащая смесь всех трех генотипов популяция подвергнется воздействию травоядных, то следует ожидать резкого повышения смертности генотипа АА и в значительно меньшей степени - генотипа Аа. Но очевидно также, что по мере потребления генотипа АА будет ослабляться внутривидовая конкуренция и повысятся шансы на выживание у генотипов Аа и аа. Увеличение доли этих генотипов будет продолжаться до тех пор, пока не ослабнет пресс травоядных (что неизбежно, поскольку доля съедобных растений будет снижаться). В условиях же ослабления выедания сможет утвердиться, а позднее занята и доминирующее положение генотип АА. Но как только это произойдет, сразу усилится пресс травоядных, и весь цикл начнется сначала. Математическая модель этого процесса показывает, что колебания долей разных генотипов действительно возникают, и вскоре они затухают, и соотношение генотипов устанавливается на некотором постоянном уровне, соответствующем заданной интенсивности выедания. Таким образом, важнейшим условием регуляции численности популяции оказывается разнокачественность составляющих ее особей. Примером крайней выраженности такой разнокачественности может быть явление «фазовой изменчивости» у нескольких видов саранчи (Schisfocerca gregaria, Locusta migratoria, Nomodacri septemfasciata и некоторых других), стаи которых время от времени совершают опустошительные налеты на посевы сельскохозяйственных культур в Африке и Азии. Хотя налеты саранчи известны с незапамятных времен, конкретные биологические механизмы, определяющие процессы увеличения численности и миграции, начали изучаться только в 20-30-е гг. нашего столетия, причем многие моменты и по сей день остаются неясными. Известно, однако, что у всех перечисленных видов имеются две сменяющие друг друга формы (фазы) - одиночная и стадная, которые настолько сильно различаются (как морфологически, так и физиологически), что ранее их относили к разный видам. По сравнению с одиночной фазой стадная характеризуется более яркой окраской, несколько другими пропорциями тела, высокой двигательной активностью и некоторым особенностями поведения, прежде всего стремлением собираться в стаи. Кроме того, у стадной фазы вылупляющиеся из яиц молодые особи (нимфы) лучше обеспечены запасами воды и питательных веществ; видимо, поэтому они отличаются более высокой выживаемостью и более быстрым развитием. Плодовитости стадной фазы меньше, чем одиночной, но яйца более крупные. В периоды между опустошительными нашествиями мигрирующие виды саранчовых, представленные одиночной фазой, встречаются в небольших количествах на довольно ограниченной территории. Подробности жизни одиночной фазы саранчовых изучены явно недостаточно, но известно, что местообитания их представляют собой открытые, поросшие редкой травянистой растительностью пространства (обязательны участки голой земли, в которую саранча откладывает яйца), подверженные чередованию засушливых и дождливых периодов. Основная часть когорты гибнет на стадии нимф, но в некоторые годы (которым предшествуют длительные засушливые периоды) выживаемость нимф резко повышается, и численность саранчи за одно поколение может увеличиться в 100 раз. Как это ни удивительно, но наиболее высокая выживаемость нимф достигается в те годы, когда растительности, служащей им пищей, бывает мало. Видимо, все дело в том, что скудная после сильной засухи растительность гораздо богаче питательными веществами (в частности, азотом) по сравнению с той, что обильно развивается в условиях достаточной обеспеченности влагой. Таким образом, не исключено, что именно определенное сочетание климатических условий служит первым толчком к будущей массовой миграции, поскольку приводит к повышению локальной плотности популяции, представленной изначально только особями одиночной фазы. Затем вследствие участившихся контактов между особями начинается превращение одиночной фазы в стадную. А поскольку особи стадной фазы быстрее размножаются, а главное, имеют ярко выраженную тенденцию собираться в группы, процесс образования стаи идет очень быстро, с нарастающей скоростью. Мигрирующие по ветру стаи могут переноситься на громадные расстояния (так, например, область, в пределах которой встречаются стаи N. septemfasciata, в 1500 раз превышает по площади область постоянного обитания одиночной фазы). Биологический смысл образования стадной фазы и миграции, по-видимому, в том, чтобы избежать риска вымирания в крайне нестабильной среде и найти новое подходящее для размножения место. Если во время миграции такое место найдено, размер стаи может увеличиваться и достигать громадных размеров. Так, стая Schistocerca gregaria, совершившая налет в Сомали в 1957 г., состояла из 1,6 х 1010 особей, и масса ее достигала 50 тыс. т. Если учесть, что за день одна саранча съедает столько, сколько весит сама, то нетрудно представить себе колоссальные размеры бедствий. Общие принципы популяционного гомеостаза То же можно сказать и о таких особенностях популяций, как средние размеры индивидуальных (или стадных) территорий, степень подвижности животных и т. п. Упоминавшаяся в гл. 10 сезонная динамика пространственной структуры у рада видов — пример устойчивого приспособления к закономерно повторяющейся смене условий жизни популяции. Характерным свойством популяции может быть и средний уровень плодовитости. Адаптивный характер этого показателя может проявляться в увеличении плодовитости в популяциях, занимающих пессимальные местообитания, что компенсирует повышенную смертность в этих условиях. Механизмы формирования и закрепления уровня стабилизации наиболее генеральных популяционных свойств основываются в первую очередь на генетических (связанных с определенной степенью изоляции) и микроэволюционных процессах. Изменчивость, динамичность конкретных условий жизни вызывают формирование лабильных функциональных адаптаций, действующих на данном уровне стабилизации популяционных функций и поддерживающих этот уровень. Механизмы таких адаптаций основаны на эколого-физиологических процессах. При всем многообразии конкретных проявлений лабильные адаптации популяционного уровня действуют по единому принципу обратной связи: в ответ на внешние или внутрипопуляционные изменения они вызывают адекватные сдвиги во внугрипопуляционных отношениях, продолжающиеся до тех пор, пока не восстановятся «уравновешенные» отношения между популяцией и средой. Именно лабильные авторегуляторные процессы придают определенную устойчивость популяционным системам на фоне изменчивых условий их жизни. В том случае, когда изменения среды выходят за пределы адаптированного диапазона, биологически выгодной оказывается смена уровня стабилизации системы. В этом случае на основе внутри-популяционной разнокачественности начинаются процессы отбора, ведущие к перестройке наиболее генерализованных качеств популяции в соответствии с изменившимися средними условиями ее жизни. Таким образом, гомеостатические механизмы популяционного уровня лежат в основе того, что в относительно стабильных условиях популяция функционирует как устойчивая форма существования вида, а в условиях направленного изменения среды представляет собой исходную элементарную единицу эволюционного процесса. Гомеостазирование популяционных функций связано с такими показателями, как численность и плотность населения. Это положение исходно предусматривает динамический характер основных популяционных параметров: Как численность популяции, так и ее выражение, отнесенное к единице пространства, не остается одинаковым во времени. Свойственные популяции процессы постоянного воспроизведения сопровождаются столь же постоянным отмиранием особей. В силу многих причин не остается неизменной и возможность заселения отдельных элементов ландшафта: «емкость угодий» меняется в сезонном и многолетнем масштабе, что определяет динамику параметров плотности населения даже при постоянном уровне репродукции. В конкретных популяциях, локализованных в определенных границах пространства, постоянно происходят процессы притока особей извне и выселения определенной части населения за пределы популяции. Все эти процессы детерминируют пульсирующий, динамический характер популяции как системы, составленной множеством отдельных организмов. Будучи представителями одного вида, организмы в составе популяции несут рад общих признаков, определяющих биологическую специфику популяции в целом и ее место в функционировании глобальных систем биологического круговорота. В то же время составляющие популяцию особи отличаются друг от друга по полу, возрасту, генетическим особенностям и месту в функциональной структуре популяции. На этой основе формируются такие фундаментальные популяционные процессы, как типы воспроизводства и смертности, масштабы и формы пространственной дисперсии, общий уровень биологической активности и др.
40. Основы биоценологии. Принципы взаимосвязи биоценоза и биотопа. После разносторонних исследований к 30-м годам XX столетия определились основные теоретические представления в области биоценологии: о границах и структуре биоценозов, степени устойчивости, возможности саморегуляции этих систем. Углублялись исследования типов взаимосвязей организмов, лежащих в основе существования биоценозов. Проблему взаимодействия живых организмов с неживой природой подробно разработал В. И. Вернадский в 1926 году, подготовив условия для понятия единого целого биологических организмов с физической средой их обитания. Часть экологии, которая исследует закономерности сложения сообществ и совместной жизни в них живых организмов, называется синэкологией, или биоценологией. Объектами экологических исследований могут быть отдельные виды (аутоэкология), популяции вида (популяционная экология), сообщества организмов (синэкология, или биоценология, геоботаника), биосфера (глобальная экология). При изучении таксономических групп выделяют экологию растений, грибов, насекомых, рыб, птиц, млекопитающих и т.д. Недавно сформировалось особое направление — экология человека. Однако в этой научной дисциплине важнейшее место занимают социальные проблемы и соответственно применяют особые методы исследований, поэтому она не может быть отнесена только к биологическим наукам. Специфическая задача экологии состоит в изучении живой природы на уровне экологических систем. Соответственно с этим основным и ведущим ее разделом следует считать синэкологию, или биоценологию, т. е. учение о сообществах растений, животных и микроорганизмов в их взаимодействии друг с другом и с неорганической средой обитания. В настоящее время биоценология переросла в науку об экосистемах, которую применительно к биоценозам суши обычно называют биогеоценологией. Общая экология классифицируется по уровням организации надорганизменных систем. Популяционная экология изучает популяции — совокупности особей одного вида, объединяемых общей территорией и генофондом. Экология сообществ (биоценология) исследует структуру и динамику природных сообществ (ценозов) — совокупностей совместно обитающих популяций разных видов. Биогеоценология — раздел общей экологии, изучающий экосистемы (биоценозы) Менее наглядна принадлежность к собственно антропогенным ландшафтам возделанных полей, культурных пастбищ, садов.[ В развитие общей экологии значительный вклад внес Д. Н. Кашкаров (1878— 1941). Ему принадлежат такие книги, как «Среда и общество», «Жизнь пустыни». Он является автором первого в нашей стране учебника по основам экологии животных (1938). По инициативе Д. Н. Кашкарова регулярно издавался сборник «Вопросы экологии и биоценологии». Наиболее четкое оформление экологии как самостоятельной научной дисциплины связано с опубликованием в 1895 г. «Экологической растительной географии» Е. Варминга. В первой половине XX в. достигла расцвета биоэкология. Решающее значение имел переход от экологии отдельного организма — аутэкологии к изучению популяций и многовидовых природных сообществ растений и животных — синэкологии и биоценологии. Основы математической экологии были заложены работами А. Лотка (1925) и В. Волътерра (1926). Заметный вклад в экологию внесен трэдами Н.И. Калабухова (1946), АД. В развитии теоретической экологии существенную роль сыграли работы Н.В. Тимофеева-Ресовского (1968) и A.A. Любищева (1972). Различают два основных типа биоценозов: естественный и антропогенный. Первый тип - это биотическое сообщество, в функционировании которого отсутствует влияние деятельности человека. Второй тип (или агробиоценоз) - это неустойчивая, искусственно созданная и регулярно поддерживаемая человеком экосистема культурных полей (поля, искусственные пастбища, сады, виноградники и т.п.). Кт.т0. ценоз является объектом изучения биоценологии -науки, которая исследует закономерности жизни организмов в природных сообществах, их популяционную структуру, потоки энергии и круговорот веществ. Произошло и смешение понятий «сообщество» и «биоценоз». Первое объединение может состоять из одних продуцентов (фитоценоз), кон-сументов (зооценоз) или микроорганизмов (микробиоценоз). Биоценоз же в классическом понимании — системно-функциональная совокупность продуцентов, консументов и редуцентов, т. е. экологически многокомпонентное образование (таков даже биоценоз мышиной норы или болотной кочки). Видимо, термин «синэкология» целесообразно сохранить за экологией сообществ, а экологию биоценозов называть биоценологией. Учение о биосфере — биосферология, а учение о среде формирования биосферы — глобальная экология, или экосферология. В связи с проблемой загрязнения среды и преобразования ее человеком возникли новые комплексные отрасли науки, называемые фитогигиеной и зоогигиеной, являющиеся частью общей и сравнительной гигиены. Содержание фитогигиены — всестороннее исследование закономерностей обеспечения здоровья растительных организмов с целью его сохранения, улучшения и искусственной регуляции. Научной базой зоо- и фитогигиены считают, в частности, такие новые отрасли науки, как техногенная фитопатология, промышленная ботаника, экологическая токсикология, индустриальная биоценология и др. Принцип системной организации вполне применим и к живой природе. Растения и животных, в том числе сельскохозяйственных, изучают на молекулярном, клеточном, тканевом, органном, организменном, популяционном, биоценозном и биогеоценозном (экосистемном) уровнях. Уже в 30-е годы в экологических работах наряду с абиотическими и биотическими стали выделять особый фактор, воздействующий на тот или иной вид через изменения как физико-химических характеристик, так и состояния биологических компонентов среды. Это — воздействие на природные условия человеческой деятельности, почему и был этот фактор назван антропогенным. Исследования биотических взаимоотношений, прослеживая взаимосвязи отдельных видов друг с другом, естественно развивались в изучение систем взимосвязанных организмов, или сообществ растений, животных, грибов, микроорганизмов. Такие сообщества, или биоценозы, стали предметом изучения биоценологии, или синэкологии — раздела экологии, занимающегося взаимодействиями в сообществах живых организмов. Исследования сложных биоценозов показывали неразрывность абиотических и биотических условий существования сообществ живых организмов на конкретной территории, что отразилось в изменении названия этого раздела экологии, который чаще и правильнее называют биогеоценологией. В развитие этой ветви экологии большой вклад внесла школа академика В. П. Сукачева. Данная задача давно привлекает внимание и лесоводов, и деятелей охотничьего хозяйства Западной Европы в том случае, когда они не представлены в одном лице. Обычно осуществляют меры защиты насаждений и охотхозяйственные мероприятия разного рода: подкормку, регулирование численности и структуры популяций и т. д. В первую очередь, это связано с дефицитом в лесах природных кормов. В лесном хозяйстве, как правило, запросы и интересы охотничьего хозяйства не учитываются. Обычно вполне отчетливо представляют себе, каким должен быть лес для наиболее эффективного комплексного его использования, мало того, одновременно в интересах рационального лесоводства. Но далее этого дело не идет. Вот уже не один десяток лет, как принцип «Оауег уа1(Га» на практике не реализуется. Причина в неизбежном при этом некотором снижении доходности от эксплуатации древесины. Здесь коммерческий интерес сегодняшнего дня не проникает глубоко в экономический анализ данной проблемы. Как отмечают некоторые крупные специалисты, дичеразведение ошибочно пытаются строить на принципах животноводства, выращивая и выкармливая копытных зверей для достижения ими определенных желаемых кондиций, без учета основ биоценологии и экологии. Биоценоз и биотоп невозможно оторвать друг от друга, об этом свидетельствует ряд принципов их взаимосвязи: 1. Принцип разнообразия (А. Тинеман): чем разнообразнее условия биотопа, тем больше видов в биоценозе. Примером проявления этого принципа может служить тропический лес. Здесь в условиях крайнего разнообразия условий среды жизни в биоценозы входит огромное число видов и трудно встретить место, где бы рядом росли два растения одного вида. 2. Принцип отклонения условий (А. Тинеман): чем выше отклонения условий биотопа от нормы, тем беднее видами и специфичнее биоценоз, а численность особей отдельных составляющих его видов выше. Этот принцип проявляется в экстремальных биотопах, например местах интенсивного загрязнения среды. В них мало видов, но число особей в них обычно велико, может иметь место даже вспышка массового размножения организмов. 3. Принцип плавности изменения среды (Г.М. Франц): чем более плавно изменяются условия среды в биотопе и чем дольше он остается неизменным, тем богаче видами биоценоз и тем более он уравновешен и стабилен. Практическое значение в том, что, чем больше и быстрее происходит преобразование природы и биотопов, тем труднее видам успеть приспособиться к этому преобразованию, а, следовательно, биоценозы ими обедняются.
41. Основные типы взаимоотношений видов в биоценозе. Правило конкурентного исключения. Типы взаимоотношений организмов в биоценозе
Экологическая ниша Экологическая ниша - место организма в природе и весь образ его жизнедеятельности, жизненный статус, включающий отношение к факторам среды, видам пищи, времени и способу питания, местам размножения и укрытий и т.д. Правило конкурентного исключения (закон Гаузе) Если два вида со сходными требованиями к среде (отношение к питанию, поведению, местам размножения, и т.д.) вступают в конкурентные отношения, то один должен либо погибнуть, либо изменить свой образ жизни и занять новую нишу.
42. Понятие об экологической нише. Механизм образования экологических ниш. Экологи́ческая ни́ша — место, занимаемое видом в биоценозе, включающее комплекс его биоценотических связей и требований к факторам среды. Термин введён в 1914 году Дж. Гриннеллом и в 1927 году Чарльзом Элтоном. В настоящее время определение Гриннелла принято называть пространственной
|