Закон Стокса: спектр люминесценции сдвинут в длинноволновую область относительно спектра поглощения того же соединения.
На рис. 24.10 приведены спектры поглощения и флуоресценции антрацена. Причина сдвига спектров заключается в том, что электронный переход при поглощении происходит с нижнего колебательного уровня основного состояния на любые колебательные уровни возбужденных электронных уровней (рис. 23.16). При этом часть энергии возбуждения молекул переходит в тепло (направленные вниз волнистые стрелки), и они оказываются на нижнем колебательном уровне нижнего возбужденного электронного состояния. Излучение происходит только с такого уровня на любые колебательные подуровни основного состояния. Длина стрелок, изображающих поглощательные и излучательные переходы в молекуле, пропорциональна энергии поглощенных или излученных фотонов. Из рис. 23.16 видно, что энергия поглощенных квантов выше, чем излученных (направленные вниз прямые стрелки короче, чем направленные вверх) — это отражает превращение части энергии возбужденного состояния молекулы в тепловую энергию окружающих молекул:
Из рис. 23.16 видно, что энергия поглощенных квантов выше, чем излученных (направленные вниз прямые стрелки короче, чем направленные вверх) — это отражает превращение части энергии возбужденного состояния молекулы в тепловую энергию окружающих молекул. Форма спектра люминесценции (правило Каши) и квантовый выход люминесценции (закон Вавилова) не зависят от длины волны возбуждения люминесценции. Причина этого в том, что излучение люминесценции происходит всегда с нижнего колебательного уровня нижнего возбужденного электронного состояния, независимо от того, на какой уровень молекула была возбуждена после поглощения. Спектр фотолюминесценции по форме совпадает со спектрами других типов люминесценции данного вида молекул, т. е. форма спектра люминесценции не зависит от способа возбуждения молекулы. Спектральное положение полосы флуоресценции (как и длинноволновой полосы поглощения) зависит от длины системы сопряженных двойных связей l: чем больше l, тем большей длине волны соответствует максимум флуоресценции lmах. Кроме того, на положение lmах (а также на квантовый выход) флуоресценции некоторых молекул могут сильно влиять свойства окружающей среды. Так, например, для диметиламинохалкона в гептане (диэлектрическая проницаемость гептана e = 1,9) lmах = 436 нм, а в воде (e = 81) lmах = 560 нм. Чувствительность параметров флуоресценции молекул к особенностям среды (диэлектрической проницаемости, вязкости и др.), в которой они оказались, используется в биологии и медицине. Если флуоресцирующие молекулы вводятся в исследуемую систему как индивидуальные вещества, которые затем сами в соответствии со своими свойствами распределяются в ней, то такие молекулы называют флуоресцентными зондами. Примером применения флуоресцентных зондов может служить внутривенное введение пациентам флуоресцеина. Этот краситель не токсичен, обладает очень высоким квантовым выходом флуоресценции (около 90%). Он с током крови разносится по всему организму и диффундирует в дерму и эпидермис. Флуоресцеин возбуждают невидимым глазом ближним УФ-светом (365 нм) и наблюдают его зеленую флуоресценцию. По скорости появления флуоресценции в поверхностных тканях судят об участках тела с пониженным кровообращением, в них флуоресцеин появляется позже, чем в участках тела с нормальным кровообращением.
В других случаях флуоресцирующие красители ковалентно связывают с какими-либо молекулами и затем вводят их в исследуемую систему. Такие молекулы называют флуоресцентными метками. Примером может служить использование флуоресцентно-меченых антител. Если добавить такие антитела к суспензии смеси клеток, то они связываются только с теми из них, на поверхности которых находятся специфические к данному антителу антигены. Возникает яркая флуоресценция определенных клонов клеток, хорошо видная в люминесцентных микроскопах, в которых в отличие от обычных источников света, как правило, используют ртутные лампы высокого и сверхвысокого давлений и применяют два светофильтра. Один из них, расположенный перед конденсором, выделяет область спектра источника света, которая вызывает люминесценцию объекта; другой, находящийся между объективом и окуляром, выделяет свет люминесценции. Подсчет таким образом меченных флуоресцирующих клеток используется в иммунологических исследованиях крови. На основе фотолюминесценции созданы источники света, спектр которых больше соответствует дневному свету, чем у ламп накаливания. Это имеет значение как для производственных целей, так и для гигиенических. В таких люминесцентных лампах, называемых лампами дневного света, происходит электрический разряд в парах ртути при низком давлении (электролюминесценция). На внутренней поверхности лампы, сделанной из обычного стекла (рис. 24.11), нанесен тонкий слой люминофора, который фотолюминесцирует под воздействием излучения паров ртути.
Изменяя состав люминофора, можно подобрать наиболее подходящий спектр фотолюминесценции. На рис. -24.12 показан один из возможных спектров, интенсивные линии соответствуют спектру паров ртути, излучение которых частично проходит через люминофор.
§ 24.7. Хемилюминесценция Хемилюминесценцией называют свечение, сопровождающее химические реакции. Наличие такого свечения означает, что энергия, которая выделяется на одной из стадий химического процесса, протекающего в системе, оказывается достаточной для образования одного из продуктов реакции в электронно-возбужденном состоянии (обозначается звездочкой): А + В -> Р* + другие продукты Р* -> Р + hv (хемилюминесценция) Среди огромного разнообразия химических реакций лишь единичные сопровождаются излучением в видимой или ультрафиолетовой областях спектра, которое удается измерить. Наиболее изучены следующие типы процессов, сопровождающихся свечением. Биолюминесценция — яркое, видимое простым глазом свечение некоторых бактерий, простейших, ракообразных, червей, моллюсков, рыб. Хорошо известна биолюминесценция светляков. Биолюминесценция обусловлена определенными ферментативными реакциями. Хемилюминесценция в системах, содержащих активные формы кислорода: пероксид водорода, супероксидный и гидроксидный радикалы, синглетный (электронно-возбужденный) кислород. Хемилюминесценция сопровождает реакции цепного (свободнорадикального, пероксидного) окисления органических соединений. Основной вклад в слабую хемилюминесценцию (раньше ее называли сверхслабым свечением) тканей, гомогенатов, растворов и суспензий полиненасыщенных жирных кислот дает свечение, сопровождающее пероксидное окисление липидов. Хемилюминесценция при реакциях свободных радикалов, образовавшихся при действии физических факторов подразделяется на следующие виды: радиохемилюминесценция (при воздействии ионизирующего излучения); фотохемилюминесценция (после облучения видимым или УФ-светом); электрохемилюми-несценция (при пропускании электрического тока через растворы); сонолюминесценция (при воздействии ультразвука); трибо-люминесценция (при перемешивании растворов) и др. При облучении (или освещении) веществ в твердой фазе (как частный случай — при облучении биологических объектов, суспензий или растворов, замороженных жидким азотом) подвижность свободных радикалов резко ограничивается. Их реакции и сопутствующая хемилюминесценция активируются при нагревании и размягчении среды. Такое свечение называют термолюминесценцией.
Измерение характеристик хемилю-минесценции — удобный метод контроля за течением химических реакций, сопровождающихся свечением, и, следовательно, может использоваться для изучения кинетики и механизма реакций. Регистрация хемилю-минесценции в ряде случаев является наиболее чувствительным методом обнаружения свободных радикалов. На рис. 24.13 показана хемилюминесценция плазмы крови при выведении в нее ионов двухвалентного железа. Железо инициирует пероксидное окисление липидов. Кванты хемилюминесценции излучаются триплетными возбужденными кетонами, образующимися при рекомбинации пероксид-ных свободных радикалов (обозначены жирными точками)
Хемилюминесценция фагоцитирующих клеток крови при добавлении к ним различных образцов промышленной пыли значительно различается: по характеру кривых свечения клеток можно оценивать токсичность пыли. Токсичная пыль активирует ферментативную систему клеток, вырабатывающую супероксидные О*2 и гидроксильные *ОН радикалы и другие активные формы кислорода. Активность фагоцитирующих клеток также резко меняется при ишемической болезни сердца, инфаркте и других заболеваниях, поэтому хемилюминесценция этих клеток используется как диагностический метод.
§ 24.8. Лазеры и их применение в медицине Несмотря на общую природу световых и радиоволн, многие годы оптика и радиоэлектроника развивались самостоятельно, независимо друг от друга. Казалось, что источники света — возбужденные частицы и генераторы радиоволн — имеют мало общего. Лишь с середины XX столетия появились работы по созданию молекулярных усилителей и генераторов радиоволн, которые положили начало новой самостоятельной области физики — квантовой электронике. Квантовая электроника изучает методы усиления и генерации электромагнитных колебаний с использованием вынужденного излучения квантовых систем. Достижения в этой области знаний находят все большее применение в науке и технике.
Ознакомимся с некоторыми явлениями, лежащими в основе квантовой электроники и работы оптических квантовых генераторов — лазеров. Создание лазеров оказалось возможным в результате реализации трех фундаментальных физических идей: вынужденного излучения (1), создания термодинамически неравновесной инверсной населенности энергетических уровней атомов (2) и использования положительной обратной связи (3). Возбужденные молекулы (атомы) способны излучать фотоны люминесценции (см. § 24.5—24.6). Такое излучение является спонтанным процессом (рис. 24.14, а). Оно случайно и хаотично по времени, частоте (могут быть переходы между разными уровнями), по направлению распространения и поляризации. Другое излучение — вынужденное, или индуцированное (рис. 24.14, б) — возникает при взаимодействии фотона с возбужденной молекулой, если энергия фотона равна разности соответствующих уровней энергии (23.31). При вынужденном (индуцированном) излучении число переходов, совершаемых в секунду, зависит от числа фотонов, попадающих в вещество за это же время, т. е. от интенсивности света, а также от числа возбужденных молекул. Другими словами, число вынужденных переходов будет тем больше, чем выше населенность соответствующих возбужденных энергетических состояний. Индуцированное излучение тождественно падающему во всех отношениях, в том числе и по фазе, поэтому можно говорить о когерентном усилении электромагнитной волны, что используется в качестве первой основополагающей идеи в принципах лазерной генерации.
В обычных условиях вынужденное излучение маловероятно. На рис. 24.15 показано «заселение» молекулами энергетических уровней, описываемое распределением Больцмана (24.24). На рисунке «длина» каждого уровня пропорциональна числу частиц, имеющих соответствующую энергию (Ео — основное состояние, Е 1, Е2,..., — возбужденные состояния, N — общее число частиц, Ni — число частиц на i-ом уровне энергии). Видно, что при «низких» температурах количество возбужденных молекул чрезвычайно мало, при повышении температуры оно увеличивается, при «высокой» температуре практически все энергетические уровни будут заселены одинаково. В любом случае, когда система находится в тепловом равновесии с окружающей средой (наиболее часто встречающаяся ситуация), большая часть молекул находится в основном состоянии. Поэтому фотоны будут сталкиваться, главным образом, с невозбужденными молекулами и будет происходить поглощение света. Для отдельной частицы равновероятны вынужденное поглощение, если частица находится в основном состоянии (рис. 24.14, в), и вынужденное излучение, если частица возбуждена (рис. 24.14, б). Поэтому даже если число возбужденных частиц в веществе равно числу невозбужденных («высокая» температура на рис. 24.15), усиления падающей электромагнитной волны не будет. На самом деле в обычном состоянии вещества («низкая» температура на рис. 24.15) условия для усиления волны не выполняются, т. е. волна при прохождении среды поглощается. Вторая идея, реализуемая при создании лазеров, заключается в создании термодинамически неравновесных систем, в которых, вопреки закону Больцмана, на более высоком уровне находится больше частиц, чем на более низком. Состояние среды, в котором хотя бы для двух энергетических уровней оказывается, что число частиц с большей энергией превосходит число частиц с меньшей энергией, называется состоянием с инверсной населенностью уровней, а среда — активной. Именно активная среда, в которой фотоны взаимодействуют с возбужденными атомами, вызывая вынужденные переходы на более низкий уровень с испусканием квантов индуцированного (вынужденного) излучения, является рабочим веществом лазера. Состояние с инверсной населенностью уровней формально получается из распределения Больцмана для Т < 0 К, поэтому иногда называется состоянием с «отрицательной» температурой. По мере распространения света в активной среде интенсивность его возрастает, имеет место явление, обратное поглощению, т. е. усиление света. Это означает, что в законе Бугера (24.3) kl < 0, поэтому инверсная населенность соответствует среде с отрицательным показателем поглощения. Состояние с инверсной населенностью можно создать, отбирая частицы с меньшей энергией или специально возбуждая частицы, например, светом или электрическим разрядом. Само по себе состояние с отрицательной температурой долго не существует. Третья идея, используемая в принципах лазерной генерации, возникла в радиофизике и заключается в использовании положительной обратной связи. При ее осуществлении часть генерируемого вынужденного излучения остается внутри рабочего вещества и вызывает вынужденное излучение все новыми и новыми возбужденными атомами. Для реализации такого процесса активную среду помещают в оптический резонатор, состоящий обычно из двух зеркал, подобранных так, чтобы возникающее в нем излучение многократно проходило через активную среду, превращая ее в генератор когерентного вынужденного излучения. Первый такой генератор в диапазоне СВЧ (мазер) был сконструирован в 1955 г. независимо советскими учеными Н. Г. Басовым и А. М. Прохоровым и американскими — Ч. Таунсом и др.1. Так как работа этого прибора была основана на вынужденном излучении молекул аммиака, то генератор был назван молекулярным. В 1960 г. был создан первый квантовый генератор видимого диапазона излучения — лазер2 с кристаллом рубина в качестве рабочего вещества (активной среды). В том же году был создан газовый гелий-неоновый лазер. Все огромное многообразие созданных в настоящее время лазеров можно классифицировать по видам рабочего вещества: различают газовые, жидкостные, полупроводниковые и твердотельные лазеры. В зависимости от типа лазера энергия для создания инверсной населенности сообщается разными способами: возбуждение очень интенсивным светом — «оптическая накачка», электрическим газовым разрядом, в полупроводниковых лазерах — электрическим током. По характеру свечения лазеры подразделяют на импульсные и непрерывные. Рассмотрим принцип работы твердотельного рубинового лазера. Рубин — это кристалл окиси алюминия А12О3, содержащий в виде примеси примерно 0,05% ионов хрома Сг3+. Возбуждение ионов хрома осуществляют методом оптической накачки с помощью импульсных источников света большой мощности.
В одной из конструкций применяют трубчатый отражатель, имеющий в сечении форму эллипса. Внутри отражателя помещены прямая ксеноновая импульсная лампа и рубиновый стержень, расположенные вдоль линий, проходящих через фокусы эллипса (рис. 24.16). Внутренняя поверхность алюминиевого отражателя хорошо отполирована или посеребрена. Основное свойство эллиптического отражателя заключается в том, что свет, вышедший из одного его фокуса (ксеноновой лампы) и отраженный от стенок, попадает в другой фокус отражателя (рубиновый стержень).
Рубиновый лазер работает по трехуровневой схеме (рис. 24.17, а). В результате оптической накачки ионы хрома переходят с основного уровня 1в короткоживущее возбужденное состояние З3. Затем происходит безызлучательный переход в долгоживущее (метастабильное) состояние 2, с которого вероятность спонтанного излучательного перехода относительно мала. Поэтому происходит накопление возбужденных ионов в состоянии 2 и создается инверсная населенность между уровнями 1 и 2. В обычных условиях переход со 2-го на 1-й уровень происходит спонтанно и сопровождается люминесценцией с длиной волны 694,3 нм. В резонаторе лазера есть два зеркала (см. рис. 24.16), одно из которых имеет коэффициент отражения интенсивности отраженного и падающего на зеркало света), другое зеркало полупрозрачное и пропускает часть падающего на него излучения (R < 100%).
Кванты люминесценции в зависимости от направления их движения либо вылетают из боковой поверхности рубинового стержня и теряются, либо, многократно отражаясь от зеркал, сами вызывают вынужденные переходы. Таким образом, пучок, перпендикулярный зеркалам, будет иметь наибольшее развитие и выходит наружу через полупрозрачное зеркало. Такой лазер работает в импульсном режиме. Наряду с рубиновым лазером, работающим по трехуровневой схеме, широкое распространение получили четырехуровневые схемы лазеров на ионах редкоземельных элементов (неодим, самарий и др.), внедренных в кристаллическую или стеклянную матрицы (рис. 24.17, б). В таких случаях инверсная населенность создается между двумя возбужденными уровнями: долгоживу-щим уровнем 2 и короткоживущим уровнем 2'. Очень распространенным газовым лазером является гелий-не-оновАй, возбуждение в котором возникает при электрическом разряде. Активной средой в нем служит смесь гелия и неона в соотношении 10: 1 и давлении около 150 Па. Излучающими являются атомы неона, атомы гелия играют вспомогательную роль. На рис. 24.17, в показаны энергетические уровни атомов гелия и неона. Генерация происходит при переходе между 3 и 2 уровнями неона. Для того чтобы создать между ними инверсную населенность, необходимо заселить уровень 3 и опустошить уровень 2. Заселение уровня 3 происходит с помощью атомов гелия. При электрическом разряде электронным ударом происходит возбуждение атомов гелия в долгоживущее состояние (со временем жизни около 10-3 с). Энергия этого состояния очень близка к энергии уровня 3 неона, поэтому при соударении возбужденного атома гелия с невозбужденным атомом неона происходит передача энергии, в результате чего заселяется уровень 3 неона. Для чистого неона время жизни на этом уровне мало и атомы переходят на уровни 1 или 2, реализуется больцмановское распределение. Опустошение уровня 2 неона происходит в основном за счет спонтанного перехода его атомов в основное состояние при соударениях со стенками разрядной трубки. Так обеспечивается стационарная инверсная населенность уровней 2 и 3 неона.
Зеркала резонатора делают с многослойными покрытиями, и вследствие интерференции создается необходимый коэффициент отражения для заданной длины волны. Чаще всего используются гелий-неоновые лазеры, излучающие красный свет с длиной волны 632,8 нм. Мощность таких лазеров небольшая, она не превышает 100 мВт. Применение лазеров основано на свойствах их излучения: высокая монохроматичность (Dl»0,01 нм), достаточно большая мощность, узость пучка и когерентность. Узость светового пучка и малая его расходимость позволили использовать лазеры для измерения расстояния между Землей и Луной (получаемая точность — около десятков сантиметров), скорости вращения Венеры и Меркурия и др. На когерентности лазерного излучения основано их применение в голографии (см. § 19.8). На основе гелий-неонового лазера с использованием волоконной оптики разработаны гастроскопы, которые позволяют голографически формировать объемное изображение внутренней полости желудка. Монохроматичность лазерного излучения очень удобна при возбуждении спектров комбинационного рассеяния света атомами и молекулами (см. § 24.3). Широкое применение лазеры нашли в хирургии, стоматологии, офтальмологии, дерматологии, онкологии. Биологические эффекты лазерного излучения зависят как от свойств биологического материала, так и от свойств лазерного излучения. Все лазеры, используемые в медицине, условно подразделяются на 2 вида: низкоинтенсивные (интенсивность не превышает 10 Вт/см2, чаще всего составляет около 0,1 Вт/см2) — терапевтические и высокоинтенсивные — хирургические. Интенсивность наиболее мощных лазеров может достигать 1014 Вт/см2, в медицине обычно используются лазеры с интенсивностью 102— 106 Вт/см2. Низкоинтенсивные лазеры — это такие, которые не вызывают заметного деструктивного действия на ткани непосредственно во время облучения. В видимой и ультрафиолетовой областях спектра их эффекты обусловлены фотохимическими реакциями и не отличаются от эффектов, вызываемых монохроматическим светом, полученным от обычных, некогерентных источников (см. § 24.9). В этих случаях лазеры являются просто удобными монохроматическими источниками света, обеспечивающими точную локализацию и дозированность воздействия. Примерами может служить использование света гелий-неоновых лазеров для лечения трофических язв, ишемической болезни сердца и др., а также криптоновых и др. лазеров для фотохимического повреждения опухолей в фотодинамической терапии. Качественно новые явления наблюдаются при использовании видимого или ультрафиолетового излучения высокоинтенсивных лазеров. В лабораторных фотохимических экспериментах с обычными источниками света, а также в природе при действии солнечного света обычно осуществляется однофотонное поглощение. Об этом говорится во втором законе фотохимии4,сформулированном Штарком и Эйнштейном: каждая молекула, участвующая в химической реакции, идущей под действием света, поглощает один квант излучения, который вызывает реакцию. Однофотонность поглощения, описываемая вторым законом, выполняется потому, что при обычных интенсивностях света практически невозможно одновременное попадание в молекулу, находящуюся в основном состоянии, двух фотонов. Если бы такое событие осуществилось, то выражение (23.31) приобрело бы вид:
что означало бы суммирование энергии двух фотонов для перехода молекулы из энергетического состояния Ек в состояние с энергией Et. He происходит также поглощения фотонов электронно-возбужденными молекулами, так как их время жизни мало, а обычно используемые интенсивности облучения невелики. Поэтому концентрация электронно-возбужденных молекул низка, и поглощение ими еще одного фотона чрезвычайно маловероятно. Однако если увеличить интенсивность света, то становится возможным двухфотонное поглощение. Например, облучение растворов ДНК высокоинтенсивным импульсным лазерным излучением с длиной волны около 266 нм приводило к ионизации молекул ДНК, подобной вызываемой у-излучением. Воздействие ультрафиолета с низкой интенсивностью ионизации не вызывало. Установлено, что при облучении водных растворов нуклеиновых кислот или их оснований пикосекундными (длительность импульса 30 пс) или наносекундными (10 нс) импульсами с интенсивностями выше 106 Вт/см2 приводило к электронным переходам, показанным на рис. 24.19, завершавшимся ионизацией молекул.
а при наносекундных (рис. 24.19, б) — по схеме
В обоих случаях молекулы получали энергию, превышающую энергию ионизации. Полоса поглощения ДНК располагается в ультрафиолетовой области спектра при l < 315 нм, видимый свет нуклеиновые кислоты совсем не поглощают. Однако воздействие высокоинтенсивным лазерным излучением около 532 нм переводит ДНК в электронно-возбужденное состояние за счет суммирования энергии двух фотонов (рис. 24.20). Поглощение любого излучения приводит к выделению некоторого количества энергии в виде тепла, которое рассеивается от возбужденных молекул в окружающее пространство. Инфракрасное излучение поглощается главным образом водой и вызывает в основном тепловые эффекты. Поэтому излучение высокоинтенсивных инфракрасных лазеров вызывает заметное немедленное тепловое действие на ткани. Под тепловым воздействием лазерного излучения в медицине понимают в основном испарение (резание) и коагуляцию биотканей. Это касается различных лазеров с интенсивностью от 1 до 107 Вт/см2 и с продолжительностью облучения от миллисекунд до нескольких секунд. К ним относятся, например, газовый СО2-лазер (с длиной волны 10,6 мкм), Nd:YAG-лазер (1,064 мкм) и другие. Nd:YAG-лaзep — наиболее широко используемый твердотельный четырехуровневый лазер. Генерация осуществляется на переходах ионов неодима (Nd3+), введенных в кристаллы Y3A15O12 иттрий-алюминиевого граната (YAG). Наряду с нагревом ткани происходит отвод части тепла за счет теплопроводности и тока крови. При температурах ниже 40 °С необратимые повреждения не наблюдаются. При температурах 60 °С и выше начинается денатурация белков, коагуляция тканей и некроз. При 100— 150 °С вызывается обезвоживание и обугливание, а при температурах свыше 300 °С ткань испаряется.
Когда излучение исходит от высокоинтенсивного сфокусированного лазера, количество выделяющегося тепла велико, в ткани возникает температурный градиент. В месте падения луча ткань испаряется, в прилегающих областях происходит обугливание и коагуляция (рис. 24.21). Фотоиспарение является способом послойного удаления или разрезания ткани. В результате коагуляции завариваются сосуды и останавливается кровотечение. Так сфокусированным лучом непрерывного СО2-лазера (l = 10,6 мкм) с мощностью около 2 • 103 Вт/см2 пользуются как хирургическим скальпелем для разрезания биологических тканей. Если уменьшать длительность воздействия (10-9—10-6 с) и увеличивать интенсивность (выше 106 Вт/см2), то размеры зон обугливания и коагуляции становятся пренебрежимо малыми. Такой процесс называют фотоабляцией (фотоудалением) и используют для послойного удаления ткани. Фотоабляция возникает при плотностях энергии 0,01—100 Дж/см2. При дальнейшем повышении интенсивности (1011 Вт/см2 и выше) возможен еще один процесс — «оптический пробой». Это явление заключается в том, что из-за очень высокой напряженности электрического поля лазерного излучения (сравнимой с напряженностью внутриатомных электрических полей) материя ионизуется, образуется плазма и генерируются механические ударные волны. Для оптического пробоя не требуется поглощения квантов света веществом в обычном смысле, он наблюдается также в прозрачных средах, например в воздухе.
1 В 1964 г. Н. Г. Басову, А. М. Прохорову и Ч. Таунсу за эти работы 2 2 Название «лазер» является аббревиатурой от англ. Light Amplifica 3 3 На этой редуцированной схеме представлены только те уровни и переходы между ними, которые существенны для генерации лазерного излучения. На самом деле схема электронных уровней гораздо сложнее. На схеме не отражена ширина энергетических уровней. Так, короткоживу-щий уровень 3 является широким и поглощает большую часть спектра излучения лампы накачки. 4 Фотохимия изучает химические реакции, протекающие под действием света.
§ 24.9. Фотобиологические процессы. Понятия о фотобиологии и фотомедицине Фотобиологическими называют процессы, которые начинаются с поглощения квантов света молекулами, а заканчиваются физиологической реакцией организма. К фотобиологическим процессам относятся фотосинтез, зрение, загар и эритема кожи, фотопериодизм и многие другие. Условно всякий фотобиологический процесс можно разбить на несколько стадий: 1) 1) поглощение кванта света молекулой; 2) 2) внутримолекулярные процессы размена энергии; 3) 3) межмолекулярные процессы переноса энергии электронно-возбужденного состояния (важны в некоторых фотобиологических процессах); 4) 4) первичный фотохимический акт, сопровождающийся образованием короткоживущих, нестабильных фотопродуктов, в него молекула вступает из нижнего синглетного S1 или триплетного Т1 возбужденных состояний; 5) 5) реакции нестабильных фотопродуктов, заканчивающиеся образованием стабильных продуктов; 6) 6) биохимические реакции с участием фотопродуктов; 7) 7) физиологический ответ на действие света. Первые три стадии фотобиологических процессов одинаковы для фотохимических реакций и фотолюминесценции. Поэтому законы фотохимии имеют свои аналогии с законами люминесценции (см. § 24.6). Первичный фотохимический акт заключается в Особенностью биологического действия ультрафиолетового и видимого излучения (200—750 нм)1 является ярко выраженная зависимость биологического эффекта от длины волны излучения. Бактерицидные эффекты вызываются волнами в диапазоне 200— 315 нм, покраснение (эритема) кожи наиболее эффективно вызывается излучением с длиной волн 280—315 нм, зрительный эффект — 400—750 нм (видимый диапазон), лечение желтухи новорожденных — фиолетовым светом (около 400 нм). При фотосинтезе растения и фотосинтезирующие бактерии используют весь диапазон солнечного ультрафиолетового излучения, достигающего поверхности Земли (коротковолновая граница солнечного света, проходящего через атмосферу Земли, ~ 285 нм), видимого света, и даже ближнего инфракрасного излучения (иногда до 1000 нм). Меняя длину волны, можно избирательно инициировать те или иные фотобиологические процессы2. Дело в том, что разные фотобиологические процессы начинаются с поглощения квантов света разными молекулами, в свою очередь положение полосы поглощения молекулы зависит от ее химической структуры (см. § 24.4). Важной характеристикой воздействия света на биологические объекты является спектр фотобиологического действия — зависимость биологического эффекта от длины волны действующего света. Спектры действия позволяют определить, какая область спектра наиболее эффективно вызывает биологический процесс, а также определить природу молекул, ответственных за поглощение света в данном процессе. Рассмотрим количественно начальные этапы этого процесса: поглощение света и первичную фотохимическую реакцию. По аналогии с рассуждениями § 24.1, введем понятие эффективного сечения поглощения молекулой фотона s. Отличие от вывода закона Бугера—Ламберта—Бера заключается, по крайней мере, в следующем: во-первых, будем учитывать уменьшение числа активируемых молекул, так как воздействие света вызывает их химические превращения; во-вторых, рассмотрим достаточно тонкий слой разбавленного раствора, это позволит считать интенсивность света I0 постоянной и одинаковой по всей толщине слоя раствора. Элементарное уменьшение концентрации dn молекул под действием света пропорционально: — — концентрации п молекул; — — эффективному сечению поглощения s; — — времени облучения dt; — — интенсивности света 103:
Здесь знак «-» означает уменьшение числа молекул со временем. Коэффициент (jх называют квантовым выходом фотохимической реакции. Этот коэффициент показывает, какая часть молекул, которые поглотили фотоны, вступила в фотохимическую реакцию. Разделим переменные и проинтегрируем (24.27):
где п0 — начальная концентрация молекул, a ni — концентрация молекул в момент i. Получаем или
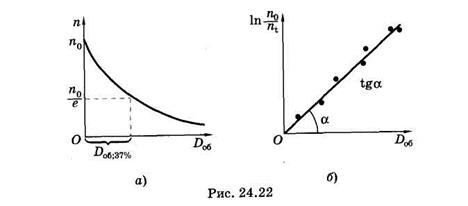
Здесь Iot = Do6 — доза облучения, a sjx = sх — эффективное сечение молекулы для фотохимического превращения, оно пропорционально вероятности такого взаимодействия фотона с молекулой, в результате которого произойдет фотохимическая реакция. Для нахождения sх строят график зависимости ln n0/ni = f(Do6) и по наклону прямой [см. (24.28)] определяют эту величину (рис. 24.22, б). Можно найти sх непосредственно на дозовой кривой фотолиза4 вещества (рис. 24.22, а)
где Do6.37% — доза облучения, при которой концентрация вещества уменьшилась в е раз, другими словами, осталось неразрушенными 37% молекул.
В фотохимии спектром действия называют зависимость s х(l). Эту зависимость можно найти, используя связь sх = sjx. Дело в том, что квантовый выход фотохимических реакций (подобно квантовому выходу флуоресценции, см. § 24.6) в растворах не зависит от длины волны действующего света, т. е. jх(l) = const. Физически это означает, что независимо от энергии возбуждения hv молекула сначала изр
|









 Основным конструктивным элементом гелий-неонового лазера (рис. 24.18) является газоразрядная трубка диаметром около 7 мм. В трубку вмонтированы электроды для создания газового разряда и возбуждения гелия. На концах трубки под углом Брюстера (§ 20.2) расположены окна, благодаря которым излучение оказывается плоскополяризованным. Плоскопараллельные зеркала резонатора монтируются вне трубки, одно из них полупрозрачное (коэффициент отражения R < 100%). Таким образом, пучок вынужденного излучения выходит наружу через полупрозрачное зеркало. Это лазер непрерывного действия. Так как уровни 2 и 3 неона обладают сложной структурой (на рис. 24.17, в не показано), то гелий-неоновый лазер может работать на 30 длинах волн в области видимого и инфракрасного диапазонов.
Основным конструктивным элементом гелий-неонового лазера (рис. 24.18) является газоразрядная трубка диаметром около 7 мм. В трубку вмонтированы электроды для создания газового разряда и возбуждения гелия. На концах трубки под углом Брюстера (§ 20.2) расположены окна, благодаря которым излучение оказывается плоскополяризованным. Плоскопараллельные зеркала резонатора монтируются вне трубки, одно из них полупрозрачное (коэффициент отражения R < 100%). Таким образом, пучок вынужденного излучения выходит наружу через полупрозрачное зеркало. Это лазер непрерывного действия. Так как уровни 2 и 3 неона обладают сложной структурой (на рис. 24.17, в не показано), то гелий-неоновый лазер может работать на 30 длинах волн в области видимого и инфракрасного диапазонов.

 При пикосекундных импульсах (рис. 24.19, а) заселение высоких электронных уровней происходило по схеме
При пикосекундных импульсах (рис. 24.19, а) заселение высоких электронных уровней происходило по схеме