Механизмы дыхания
У амфибий в отсутствие грудной клетки легкие обслуживаются рототлоточной полостью — попутно, помимо адаптации, саязанных питанием. Это гулярное дыхание, нагнетательное. Оно требует хорошо управляемой вертикальной подвижности дна ротоглоточной ции (т.е. вертикальной подижности). В результате для многих хвостатых оказалось выгодным вторично отказаться от легочного дыхания.
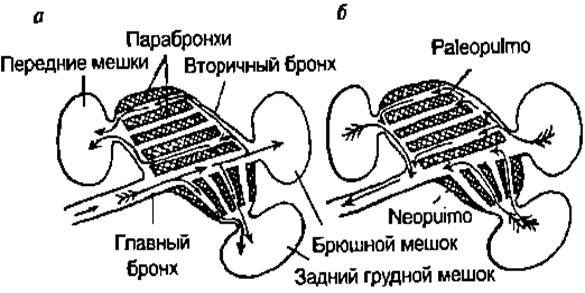
У рептилий, как и у всех остальных амниот, развито реберное, всасывательное дыхание. Грудная клетка снимает отмеченные для амфибий ограничения производительности легочного дыхания, поскольку может обслужить легкие любого строения и объема. Как и все амниоты, рептилии имеют длинную (соответственно прибавке длины шеи) трахею, армированную хрящевыми или костными кольцами. Одно из важных последствий приобретения реберного дыхания — отказ от кожного дыхания и сильное ороговение кожи. У черепах с их неподвижной грудной клеткой необходимое для выполнения вдоха увеличение внутреннего объема тела осуществляется благодаря изменению кривизны участков мягкой кожи, которая затягивает вырезки панциря н подстилается тонкими мышцами. При выдохе легкие сжимает окружающая их пленка, подтягиваемая специальными мышцами (диафрагмальными), а также охватывающая тонким слоем все внутренности поперечная мышца живота. Находясь в защитной позе, черепаха дышать не может- следовательно, длительное использование такой защиты требует устойчивости к гипоксии (как и пребывание под водой у водных черепах). У птиц вентиляция легких осуществляется только благодаря экскурсиям грудной клетки (изменениям ее объема вследствие движений грудины и ребер) как в покое, так и в полете. При вдохе воздух проходит по магистральному бронху в задние воздушные мешки (рис. 124), а часть его расходится по дорсобронхам и засасывается по парабронхам палеопульмо в передние мешки. При выдохе использованный воздух из этих мешков выходит непосредственно в магист- I ральный бронх и по нему — наружу, а из задних мешков свежий воздух проходит по дорсо- и латеробронхам к парабронхам, через них попадает в вентробронхи и оттуда — в главный бронх. Клапанов в этой системе не обнаружено, столь четкий порядок вентиляции поддерживается ее чисто гидродинамическими свойствами (при дозвуковых скоростях воздух подчиняется законам гидродинамики). В частности, при вдохе слишком низкое давление в главном бронхе (заансяшсс, по закону Бсрнушки, от скорости потока) препятствует прямому движению воздуха по вентроб-ронхам в передние легочные мешки. Именно поэтому воздух может попасть туда только окольным путем — через дорсобронхи и пара-бронхи палеопульмо.
ног о дыхания), причем в одном и том же направлении (сзади вперед). Этим достигается высокая эффективность газообмена в легких, позволяющая птицам летать на высотах до 10 км (точный замер — 9600 м; млекопитающие на такой высоте вообще не могут жить.
У млекопитающих процесс дыхания очень прост: это набирание воздуха при расширении легких и выпускание при их частичном спаде- w/и, как и у рептилий. Своеобразен лишь механизм управления изменениями объема грудной клетки при вдохе; участие в нем длинных продольных лестничных мышц, при опоре на шею полнима-юших каждое ребро, словно ручку ведра, и куполообразного мускулистого «донышка» грудной клетки — диафрагмы
ции (т.е. вертикальной подвижности). В результате для многих хвостатых оказалось выгодным вторично отказаться от легочного дыхания. У рептилий, как и у всех остальных амниот, развито реберное, всасывательное дыхание. Грудная клетка снимает отмеченные для амфибий 01раничения производительности легочного дыхания, по-1 скольку может обслужить легкие любого строения и объема. Как и все амниоты, рептилии имеют длинную (соответственно прибавке длины шеи) трахею, армированную хряшевыми или костными кольцами. Одно из важных последствий приобретения реберного дыхания — отказ от кожного дыхания и сильное ороговение кожн. У черепах с их неподвижной грудной клеткой необходимое для выполнения вдоха увеличение внутреннего объема тела осуществляется благодаря изменению кривизны участков мягкой кожи, которая затягивает вырезки панциря н подстилается тонкими мышцами. При выдохе легкие сжимает окружающая их пленка, подтягиваемая специальными мышцами (так называемыми диафрагмальными), а также охватывающая тонким слоем все внутренности поперечная мышца живота. Находясь в защитной позе, черепаха дышать не может- следовательно, длительное использование такой защиты требует устойчивости к гипоксии (как и пребывание под водой у водных черепах). У птиц вентиляция легких осуществляется только благодаря экскурсиям грудной клетки (изменениям ее объема вследствие движений грудины и ребер) как в покое, так и в полете. При вдохе воздух проходит по магистральному бронху в задние воздушные мешки (рис. 124), а часть его расходится по дорсобронхам и засасывается по парабронхам палеопульмо в передние мешки. При выдохе использованный воздух из этих мешков выходит непосредственно в магист- I ральный бронх и по нему — наружу, а из задних мешков свежий воздух проходит по дорсо- и латеробронхам к парабронхам, через них попадает в вентробронхи и оттуда — в главный бронх. Клапанов в этой системе не обнаружено, столь четкий порядок вентиляции поддерживается ее чисто гидродинамическими свойствами (при дозвуковых скоростях воздух подчиняется законам гидродинамики). В частности, при вдохе слишком низкое давление в главном бронхе (заансяшсс, по закону Бсрнушки, от скорости потока) препятствует прямому движению воздуха по вентроб-ронхам в передние легочные мешки. Именно поэтому воздух может попасть туда только окольным путем — через дорсобронхи и пара-бронхи палеопульмо. Таким образом, парабронхи палеопульмо продуваются свежим воздухом как при вдохе, так и при выдохе (отсюда и понятие двой ног о дыхания), причем в одном и том же направлении (сзади вперед)- Этим достигается высокая эффективность газообмена в легких, позволяющая птицам летать на высотах до 10 км (точный замер — 9600 м; млекопитающие на такой высоте вообще не могут жить), но создающая опасность гиперпноэ при принудительной вентиляции в покое, когда интенсивность метаболизма приблизительно в 10 раз ниже, чем в полете. Такая опасность возникает при пении (особенно у воробьиных). Обнаружено несколько механизмов временного снижения эффективности газообмена и тем самым интенсивности вымывания углекислоты. Во-первых, в легких имеется относительно менее эффективный отдел — неопульмо, парабронхи которого вентилируются при выдохе в обратном направлении и притом исполь-зованным воздухом. Во-вторых, углекислоту сберегает обширное мертвое пространство трахеи, которая дополнительно удлинена у многих птиц, об;[адаюших мощной вокализацией (например, у лебедя-кликуна в сравнении с лебедем-шипуном). Однако, как показывают расчеты, этих механизмов недостаточно. Остается предполагать, что в легких имеются сосудистые анастомозы (замыкаемые гладкой мускулатурой), которые при повышении рН крови восполняют дефицит С02, непосредственно пропуская венозную кровь из легочной артерии в легочную вену. У млекопитающих процесс дыхания очень прост: это набирание воздуха при расширении легких и выпускание при их частичном спаде- w/и, как и у рептилий. Своеобразен лишь механизм управления изменениями объема грудной клетки при вдохе; участие в нем длинных продольных лестничных мышц, при опоре на шею полнима-юших каждое ребро, словно ручку ведра, и куполообразного мускулистого «донышка» грудной клетки — диафрагмы.
рис. 124. Схема легких птиц и движения воздуха в них: а — при вдохе; 6 — при выдсхе Дыхательная система
|