Типы мышечных волокон.
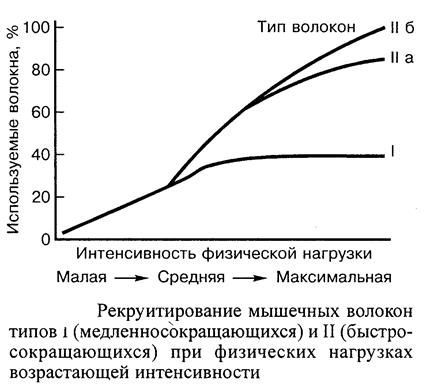
Существование различных типов волокон в скелетных мышцах достаточно очевидно и их изучение продолжается на протяжении длительного периода. Однако детальные физиологические и биохимические основы этой дифференциации, а также их функциональное значение установлены лишь в последнее время. Во многом стимулами для этих исследований явилось понимание того, что успех в спортивных соревнованиях, требующих способности спортсмена к проявлению либо максимальной силы, либо выносливости, существенно зависит от пропорционального соотношения в мышце типов волокон. Мышечные волокна, однако, чрезвычайно пластичны, и хотя их соотношение генетически детерминировано, реализация определенной тренировочной программы способна во многом повлиять на метаболический потенциал мышцы независимо от соотношения в ней типов волокон. Первоначальной основой для разделения мышечных волокон на красные, белые и промежуточные применительно к целостной мышце послужил простой визуальный осмотр. Однако основной функциональной характеристикой дифференциации типов мышечных волокон является скорость их укорочения и расслабления. Медленносокращающиеся волокна характеризуются относительно продолжительным временем, необходимым для достижения пика напряжения (около 80-100 мс для волокон мышц человека), а также длительным полупериодом расслабления. В противоположность им для достижения пика напряжения быстросокращающимися волокнами у человека требуется около 40 мс, следовательно, время их расслабления соответственно короче. Эти два типа волокон образуют отдельные группы с частичным совпадением их свойств. Поскольку для изучения механических свойств волокон доступ к ткани затруднителен, а небольшие образцы мышцы относительно легко могут быть получены с помощью пункционной биопсии, классификация типов мышечных волокон обычно базируется на биохимическом окрашивании их поперечных срезов. На этой основе волокна мышц человека обычно подразделяют на три главные группы — типы I, На и Пб, хотя возможно также дальнейшее подразделение. Аналогичное деление свойственно и мышцам животных, волокна которых классифицируются на основе прямого определения их функциональных свойств: соответственно медленносокращающиеся, быстросокращающиеся медленноутомляемые и быстросокращающиеся быстроутомляемые. Миозин различных типов мышечных волокон существует в различных молекулярных формах (изоформах), а АТФ-азная активность миофибрилл проявляет неодинаковую чувствительность к рН. Миозиновая АТФ-аза волокон типа II инактивируется при низких значениях рН (менее 4,5), тогда как АТФ-азная активность миозина в волокнах типа I остается неизменной. При значениях, превышающих рН 9, ситуация изменяется: АТФ-азная активность миозина в волокнах типа II остается стабильной, в то время как аналогичная активность в волокнах типа I снижается. Эти два отдельных подтипа типа II можно распознать путем преинкубации волокон при значениях рН, которые находятся в конце указанного диапазона: АТФ-азная активность миозина в волокнах типа Па инактивируется при рН 4,6-4,8, в то время как в волокнах типа IIб она не изменяется. Иногда путем преинкубации можно выявить и волокна типа IIв, но этот тип вряд ли стоит принимать во внимание, поскольку в мышцах человека он составляет менее 1 % всех волокон. Волокна типа IIа являются красными клетками, в которых метаболические и физиологические характеристики находятся между крайними значениями других двух типов волокон. Они содержат высокоактивную миозиновую АТФ-азу, аналогичную волокнам типа IIб, однако характеризуются окислительной способностью, более близкой к волокнам типа I. Результаты последних исследований показывают, что для большинства мышечных волокон их фактическая идентификация определяется одной и более изоформой миозина. Такая совместная экспрессия различных изоформ миозина создает предпосылки для расширения диапазона сократительных характеристик внутри определенного типа волокон (классифицируемых в соответствии с окраской гистологических образцов) и степени их частичного совпадения между типами волокон при определении их сократительных характеристик. В действительности мы продолжаем оперировать сократительными и биохимическими характеристиками, соответствующими двум крайним типам классификации, определяемым как волокна типов I и IIб. С различиями в скорости сокращения и метаболического профиля основных типов мышечных волокон отмечается дифференциация и двигательных нейронов, иннервирующих эти волокна. Так, волокна типа I иннервируются нейронами, имеющими малый диаметр и характеризующимися низкой скоростью проведения потенциала действия, а также низким порогом активации. Волокна типа II иннервируются нейронами большего диаметра, которые отличаются относительно более высокой прочностью и более высоким порогом возбудимости. Различие в порогах возбудимости двигательных нейронов связано с типом мышечных волокон, которые они иннервируют, с их рекрутированием при мышечных нагрузках, что и определяет метаболический ответ на последние. При различных видах двигательной активности проявляется определенная иерархия включения в нее двигательных единиц, в зависимости от их размера, которая, в общих чертах, соответствует волокнам типов I и II. Это свидетельствует о том, что, при легких физических нагрузках, в наибольшей степени задействованы волокна типа I, при нагрузках средней мощности — волокна типов I и II, а при более напряженной мышечной деятельности — все типы волокон включаются в работу. Все мышцы организма представляют собой смесь указанных выше трех типов волокон, хотя пропорциональное соотношение в каждой из них и в каждом из индивидуумов обнаруживает существенное различие.
Так, мышцы, отвечающие за поддержание позы тела, содержат высокий процент (обычно более 70 %) волокон типа I, функция которых состоит в поддержании пролонгированного, но относительно невысокого напряжения. Однако быстросокращающиеся волокна типа II преобладают в мышцах, выполняющих быстрые движения (например, в мышцах руки и глаза). Другие же мышцы, как, например, четырехглавая мышца бедра, имеют разное соотношение типов волокон. Их композиционный состав генетически детерминирован и не поддается значительным изменениям под влиянием тренировки. Следовательно, способность к достижению высоких спортивных результатов является в значительной степени врожденной (с учетом того, что генетический потенциал индивидуума реализуется при соответствующей методике тренировки и питания). Исследование m. vastus lateralis у марафонцев высокой квалификации показало, что эта мышца. содержит больший процент (около 80 %) волокон типа I, тогда как у элитных спринтеров обнаружился более высокий процент (около 60 %) быстросокращающихся волокон типа II Безусловно, такие исследования требуют применения методики пункционной биопсии, которая позволяет изъять около 1000 волокон в одной пробе. Относительно небольшое количество исследований по изучению распределения волокон в мышцах человека было проведено при вскрытии трупов. Региональная вариативность композиционного состава мышц проявляется незначительно, хотя и отмечается тенденция к тому, что в глубоколежащих слоях преобладают волокна типа I. Это подтверждается и данными, полученными с помощью достаточно надежного метода — пункционной биопсии. Исследования, проведенные на m. vastus lateralis у человека, свидетельствуют о том, что разница в показателях между пробами составляет не более 6 % (коэффициент вариации) по отношению к доминирующему типу волокон. Типы мышечной активности. Скелетные мышцы могут проявлять три различных типа активности: изометрический, при котором длина мышцы остается постоянной; концентрический, когда мышца укорачивается; эксцентрический, при котором длина мышцы, находящейся в активном состоянии, увеличивается. Обычно подразумевается, что мышцы развивают силу при сокращении. Однако тщательные исследования, проведенные на изолированных мышцах, свидетельствуют о том, что проявление силы не сопровождается изменением объема мышцы. Для проявления силы мышцы стремятся сократиться (т. е. уменьшить длину саркомеров), что в результате может иметь три основных исхода. В случае концентрической активности мышца выполняет механическую работу и развивает при этом определенную мощность. Если мышца проявляет активность в условиях изометрического или эксцентрического сокращения, то мощность будет поглощаться. Такие физические упражнения, выполняемые как при одном из видов активности, так и в комбинации, способны вызвать нарушение гомеостаза. Наиболее важной комбинацией может быть цикл «удлинение-сокращение», в котором эксцентрической активности непосредственно предшествует концентрическая активность. Сохранение возможности проявления высокой мощности в начале эксцентрической фазы обеспечивается энергией, запасенной в мышце за счет ее эластичности. Во время последующей, генерирующей мощность, концентрической фазы вклад восстанавливающей эластической энергии препятствует развитию утомления по сравнению со случаем чисто концентрической активности. Повторное выполнение высокомощностных эксцентрических действий может вызвать повреждение мышц и возникновение в них временных болезненных ощущений, которые обычно появляются через 6-12 ч после физической нагрузки и сохраняются несколько дней. Роль различных типов волокон в эксцентрической мышечной активности еще полностью не выяснена, хотя, скорее всего, в данном случае активируется небольшое количество волокон по сравнению с концентрическим сокращением. Имеется также несколько доказательств, что волокна типа II могут избирательно рекруитироваться во время эксцентрической активности мышц. Такая высокая нагрузка на относительно небольшое количество волокон может вызвать их локальные повреждения, связанные с воспалительным процессом и сопровождающимися отечностью и болевыми ощущениями. Доказательством того, что в мышцах действительно происходят повреждения, служит появление в крови высоких уровней внутримышечных ферментов в дни выполнения эксцентрических упражнений, а также гистологические доказательства разрушения саркомеров и Z-линий в работавших мышцах. Пластичность скелетных мышц. Скелетная мышца является чрезвычайно пластичной тканью: она обладает значительной способностью адаптироваться к различным видам активности либо к бездействию. Адаптация может проявляться в изменении размеров мышцы, композиционного состава волокон, метаболической способности, плотности капилляризации. Размер мышц и их функции с возрастом изменяются. Максимальной силы мышцы у мужчин и женщин достигают в возрасте между 20 и 30 годами. К 70 годам сила мышц снижается в среднем на 30 %. Уменьшение мышечной массы является ведущим фактором в этом процессе. Возможно также, что происходящее с возрастом снижение мышечной силы связано с уменьшением размеров волокон, в частности волокон типа II. Не исключено уменьшение при этом общего количества мышечных волокон, обусловленное утратой к старости двигательных нейронов. Иннервация мышечных волокон необходима для поддержания их существования (возможно, это связано с выработкой в нерве факторов роста), а денервация приводит к атрофии мышечных волокон и замещению их соединительной тканью. Связанное со старением снижение мышечной массы может происходить как за счет самих возрастных изменений, происходящих в организме, так и в результате снижения двигательной активности либо одновременно за счет обоих факторов. Однако ясно, что и в старом организме мышцы еще сохраняют способность адаптироваться в ответ на силовую тренировку и что значительного улучшения физиологических, структурных и физических характеристик можно достичь без реализации напряженных тренировочных программ. В молодом возрасте частота, интенсивность и продолжительность упражнений являются решающими факторами, определяющими диапазон происходящих под влиянием тренировки адаптационных изменений
|