
Загальна характеристика коренеплідних овочевих культур
До групи овочевих коренеплідних культур входять рослини з ботанічних родин — капустяних (редька, редиска, бруква, ріпа), селерових (морква, пастернак, петрушка, селера) і лободових (столові буряки). Продуктивним органом цих культур є коренеплід, який складається з власне кореня і надземних органів рослин, де відкладається запас поживних речовин. У ранньому віці ці культури мають стрижневий корінь, а коренеплоди формуються після утворення достатньо великої асиміляційної поверхні, яка дає змогу рослинам створити певний запас поживних речовин. Пізніше коренеплоди набувають типових форми, розмірів, забарвлення і будови. Розрізняють головку коренеплоду, на якій розміщуються листки, прикріплені основою черешків до головки, і в пазухах листків — бруньки; шийку коренеплоду — середню частину, на якій немає листків і бічних відгалужень кореня; власне корінь — нижню частину коренеплоду, на якій розміщені бічні корінці. Головка коренеплодів утворюється з епікотиля (надсім’ядольного коліна). Шийка коренеплоду розвивається з гіпокотиля (підсім'ядольного коліна) при його потовщенні і розростанні у висоту. Власне корінь утворюється в результаті потовщення верхньої частини кореня. У редиски, столових буряків, пастернаку з округлою формою коренеплодів більша частина продуктивного органа утворюється за рахунок розростання підсім'ядольного коліна, а тому заглиблення коренеплодів у грунт у них незначне. У сортів редиски, буряків, пастернаку, петрушки, моркви з конічною формою коренеплодів продуктивні органи утворюються за рахунок розростання власне кореня і, як правило, заглиблюються в грунт майже повністю. Сорти столових буряків за формою округлі, округло-плоскі, плоскі, за забарвленням — фіолетово-червоні або бордово-червоні з перехідними відтінками. Частіше трапляються циліндрична, конічна, овальна, округла форми коренеплодів моркви. Забарвлення коренеплодів моркви жовто-оранжеве, інтенсивно-оранжеве.. Коренеплоди петрушки видовжено-циліндричні, циліндричні, конічно-гострокінцеві. Коренеплоди пастернаку округлі, округло-плоскі, конусоподібні з поступовим звуженням донизу. Коренеплоди селери округло-плоскі, округлі з великою кількістю корінців у нижній частині коренеплоду. У редьки і редиски коренеплоди округло-плоскі, округлі, конічні й циліндричні. За забарвленням коренеплоди петрушки, селери — білі; пастернаку — кремувато-білі; редиски — білі, рожеві, червоні різних відтінків, червоні з білим кінцем; редьки — білі, чорні, фіолетові. Коренеплоди ріпи і брукви округлі або плоско-округлі, забарвлення їх біле, кремово-жовте, ясно-фіолетове. За анатомічною будовою розрізняють три типи коренеплодів: редьковий (редька, редиска, ріпа, бруква), морквяний (морква, пастернак, петрушка, селера), буряковий (столові буряки). Первинна будова коренеплодів усіх типів у фазі сім'ядольних листків однакова. Гіпокотиль зверху покритий шкірочкою, під якою знаходиться первинна флоема, камбіальне кільце і в центрі первинна ксилема. З початком утворення справжнього листка коренеплоди у різних типів розвиваються неоднаково. Коренеплід редькового типу потовщується за рахунок інтенсивного поділу клітин камбіального кільця, які всередину коренеплоду відкладають соковиті паренхімальні клітини вторинної ксилеми (деревини), а на зовнішній бік — клітини вторинної флоеми (кори). Внаслідок такого нерівномірного наростання клітин ксилеми і флоеми камбій у міру росту коренеплоду в товщину переміщується до його периферії. Флоема у сформованих великих коренеплодів редьки не перевищує 3—4 мм, а ксилема (деревинна паренхіма) займає всю внутрішню їстівну частину (рис. 1, а). У редиски їстівна і зовнішня корова частина коренеплодів. Первинна флоема тріскається вздовж, відокремлюється і замінюється шаром вторинної флоеми. Цей процес скидання первинної кори дістав назву линяння коренеплодів. Сформований коренеплід культур, що характеризуються редьковим типом будови, зверхупокритий кірковою тканиною; далі розміщений тонкий шар (2—4 мм) вторинної флоеми (кори), далі до центру — вторинна ксилема (деревина) з радіальними променями деревної паренхіми і судинних пучків, в центрі розміщена первинна ксилема, розділена первинними променями паренхіми. Морквяний тип будови коренеплодів (рис. 1, б) характеризується інтенсивним розвитком як вторинної флоеми, так і вторинної ксилеми.-У сформованого коренеплоду такого типу середня частина — це ксилема, а зовнішнє кільце — флоема. Камбіальне кільце первинного судинно-волокнистого пучка гіпокотиля після утворення першого справжнього листка протягом періоду вегетації утворює клітини вторинної ксилеми у напрямі до центра коренеплоду, а до периферії утворює клітини вторинної флоеми.
Як і у коренеплодів редькового типу, первинна флоема злущується. На периферичній частині коренеплоду розміщуються старіші клітини вторинної флоеми, які покриті тонким шаром покривних окорковілих еластичних клітин. У моркви шар покривних клітин дуже тонкий і становить лише 30—75 мк. Тому коренеплоди моркви на повітрі і особливо на сонці швидко в'януть, втрачають воду. На відміну від коренеплодів редькового типу у столових буряків потовщення відбувається за рахунок вторинної і третинної будови тканин (рис. 2). При первинній будові гіпокотиль і корінь у центрі мають первинну ксилему. Навколо неї розміщуються одношарове кільце клітин перициклу (первинний камбій) і шар первинної флоеми (кори), а зверху вони покриті шкірочкою. При потовщенні відбувається поділ клітини одношарового перициклу, утворюється у внутрішньому напрямі до центра вторинна паренхімна тканина. При цьому клітини переміщуються до периферії. З шару паренхіми, що прилягає до первинної ксилеми, утворюється вторинна ксилема. Пізніше з розвитком вторинної паренхіми перициклу, пересуваючись до периферії, вторинна ксилема розриває первинну флоему (кору) і злущує її. Так відбувається линяння гіпокотиля у буряків. При цьому на поверхні перициклу утворюється тонка вторинна флоема, ззовні покрита кірковою тканиною. Перицикл протягом усього періоду вегетації і росту коренеплоду до внутрішнього боку утворює вторинну паренхіму і до зовнішнього боку-клітини вторинної флоеми. Сформоване нове кільце камбію знову до середини утворює клітини ксилеми, де розміщуються судинно-волокнисті провідні пучки, а на зовнішній бік утворює флоему. Цей цикл повторюється багаторазово. Внаслідок третинного росту коренеплоди буряків досягають великих розмірів за рахунок потовщення кілець ксилеми із сильно розвиненою судинно-волокнистою системою. Сильно розвинена в коренеплодах ксилема знижує їх якість. Щоб запобігти надмірному розростанню коренеплодів за рахунок ксилеми, треба запобігати зрідженню посівів, не вносити високих доз органічних і мінеральних азотних добрив. Діаметр коренеплодів столових буряків має становити не більш як 10—12 см, тоді анатомічна структура їх буде дрібноклітинною з помірно розвиненою ксилемою і добре розвиненою флоемою. Забарвлення коренеплоду буде інтенсивне, без білих кілець, а вміст екстрактивних сухих речовин високий, лежкість - добра.
а — під час линяння (скидання первинної кори); б — після линяння; 1-первинна флоема; 2-вторинна флоема; 3-вторинна ксилема; 4-первинна ксилема; 5-судинно-волокнистий пучок; 6-перидерма; 7 — первинний покрив (первинна кора коренеплоду).
|
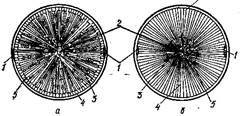 Рис. 1. Схема поперечного розрізу столових коренеплодів: а — поперечний розріз коренеплоду редьки; б — поперечний розріз коренеплоду моркви; 1 — первинний луб; 2 — первинна деревина (ксилема); 3 — вторинна деревина з широкими променями; 4 — вторинний луб; 5 — камбій.
Рис. 1. Схема поперечного розрізу столових коренеплодів: а — поперечний розріз коренеплоду редьки; б — поперечний розріз коренеплоду моркви; 1 — первинний луб; 2 — первинна деревина (ксилема); 3 — вторинна деревина з широкими променями; 4 — вторинний луб; 5 — камбій.

 Рис. 2. Схема будови коренеплодів буряків:
Рис. 2. Схема будови коренеплодів буряків:


