Спонтанный и индуцированный мутагенез
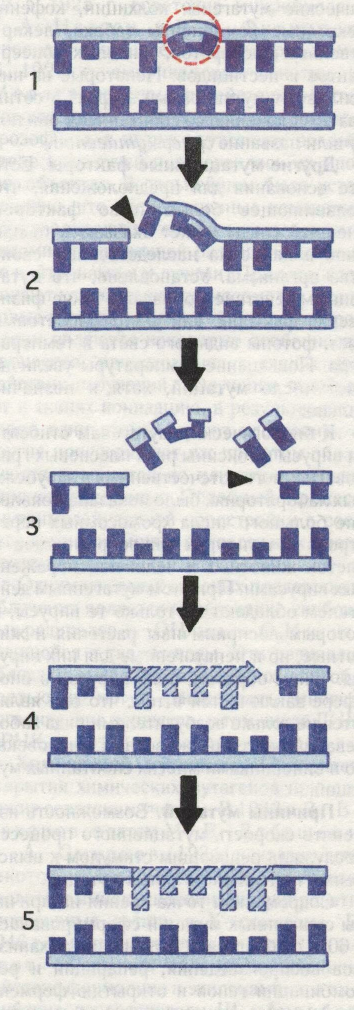
Спонтанными называют мутации, возникающие под влиянием неизвестных природных факторов, чаще всего как результат ошибок при воспроизведении генетического материала (ДНК или РНК), поскольку репликация не происходит с абсолютной точностью, а процессы репарации (исправление повреждений в молекуле ДНК) не обладают абсолютной эффективностью. Частота спонтанного мутирования у каждого вида генетически обусловлена и поддерживается на оптимальном уровне— 106—107. До 1925—1927 гг. генетики имели дело только со спонтанными мутациями. Различные попытки повысить частоту мутаций не приносили успеха. Создавалось впечатление, что мутационный процесс не зависит от условий окружающей среды. Радиационный мутагенез. Впервые повышение частоты наследственной изменчивости под влиянием внешних агентов (ионизирующая радиация) обнаружили в 1925 г. советские микробиологи Г. А. Надсон и Г. С. Филиппов, проводившие исследования с дрожжами. В 1927 г. американский генетик Г. Меллер сообщил о действии рентгеновских лучей на мутационный процесс у дрозофилы, а позже мутагенное воздействие этих лучей подтвердилось на многих объектах. В дальнейшем было установлено, что наследственные изменения обусловливаются также всеми другими видами проникающей радиации, особенно нейтронами и γ-лучами. При этом возникают как разрывы хромосом, так и точковые мутации. Причиной разрывов хромосом являются некоторые особенности процессов, происходящих в тканях при действии излучений. Излучения вызывают в тканях ионизацию, в результате которой одни атомы теряют электроны, а другие присоединяют их; при этом образуются положительно или отрицательно заряженные ионы. Подобный процесс внутримолекулярной перестройки в составе хромосом может привести к их фрагментации. Одним из самых опасных последствий облучения является образование свободных радикалов — ОН или — О2Н из находящейся в тканях воды. Эти радикалы обладают высокой реакционнной способностью и могут расщеплять многие органические вещества, в том числе ДНК и РНК. Химический мутагенез. Приоритет открытия химических мутагенов принадлежит советским ученым. В 1933 г. В. В. Сахаров,а затем М. Е. Лобашеви Ф. А. Смирнов (1934) показали, что некоторые соединения (йод, уксусная кислота, аммиак) способны индуцировать рецессивные летали в Х-хромосоме. В 1946 г. советский генетик И. А. Рапопорт обнаружил сильное мутагенное действие формалина и этиленамина, а английские исследователи Ш. Ауэрбах иДж. Робсон — азотистого иприта. Позже были открыты многие другие химические мутагены: колхицин, кофеин, некоторые компоненты табака, лекарственных препаратов, пищевых консервантов и пестицидов. Некоторые из них усиливают мутационный эффект в сотни раз по сравнению со спонтанным; они получили название супермутагенов. Другие мутационные факторы. Есть все основания для предположения, что подавляющее большинство факторов внешней среды может оказывать то или иное влияние на наследственные свойства организма. Установлено, что мутагенным действием обладают такие физические факторы, как ультрафиолетовый свет, фотоны видимого света и температура. Повышение температуры увеличивает число мутаций, хотя и незначительно. К биологическим мутагенам относятся вирусы и токсины ряда плесневых грибов. Во многих отечественных и зарубежных лабораториях было показано появление большого числа хромосомных перестроек в культурах микроорганизмов и клеток животных и человека, пораженных вирусами. При этом мутагенным действием обладают не только те вирусы, к которым восприимчивы растения и животные, но и непатогенные для них вирусы. Таким образом, роль вирусов в биосфере заключается в том, что они являются не только возбудителями ряда заболеваний растений, животных и человека, но и виновниками многих спонтанных мутаций. Причины мутаций. Возможность изменять скорость мутационного процесса послужила решающим стимулом к выяснению причин спонтанных мутаций. Современная точка зрения на причины спонтанных мутаций сформировалась в 60-е гг. благодаря выяснению механизмов воспроизведения, репарации и рекомбинации генов и открытию ферментных систем, ответственных за эти процессы. Возникла гипотеза, согласно которой генные мутации — это ошибки в работе ферментов матричного синтеза ДНК. Притягательность этой общепризнанной гипотезы заключается в том, что она позволяет дать единое объяснение причин спонтанных и индуцированных мутаций, происходящих под генетическим контролем. Были открыты гены, мутации которых могут повышать или понижать частоту как спонтанных, так и индуцированных мутаций. Мутация рассматривается как результат перестройки атомов в сложной молекуле гена. Причиной такой перестройки является попадание в ген кванта света или ионизирующей частицы или же случайные колебания атомов. Причем мутации возникают в результате процесса, длящегося во времени, а не непосредственно в момент прохождения кванта энергии или ионизирующей частицы через ген. Согласно физиологической гипотезе М. Е. Лобашева (1946), появлению мутаций должно предшествовать предмутационное состояние или потенциальное изменение, которое может быть устранено в процессе репарации либо реализуется в виде мутаций. Связь мутаций с процессами репарации в настоящее время доказана практически для всех исследованных объектов. Репарация ДНК и мутационный процесс. Под действием различных физических и химических агентов могут возникнуть повреждения ДНК в виде димеров тимина, сшивок между нитями или с белком, разрывов ДНК, появления нуклеотидов аномальной структуры и т. д. Повышенная частота этих повреждений блокирует репликацию ДНК, а тем самым и деление ядра и клетки. Оказалось, что клетка обладает различными механизмами исправления повреждений в нитях ДНК. Способность клеток всех организмов к самовосстановлению поврежденной первичной структуры ДНК получила название репарации. В настоящее время выявлены три основных механизма репарации ДНК: фотореактивация, эксцизионная репарация и пострепликативная репарация. Последние два типа называют также темповой репарацией, поскольку и та и другая не зависят от света (рис. 160). Лучше всего изучена репарация повреждений, вызванных ультрафиолетовыми лучами. При облучении ультрафиолетом между соседними пиримидиновыми основаниями одной цепи возникают димеры, чаще всего димер Т — Т, т. е. вместо водородных связей между Т и А двух комплементарных нуклеотидных цепей образуются связи Т — Т внутри одной цепи. Фотореактивация происходит при воздействии видимого света. При этом репарирующий фермент связывается с поврежденной ДНК, разделяет димер на мономеры и водородные связи Т — А между комплементарными цепями опять восстанавливаются. Если при фотореактивации исправляются повреждения, возникшие только под воздействием ультрафиолетовых лучей, то при темновой репарации — повреждения, появившиеся под влиянием других физических, химических и биологических факторов. При эксцизионной репарации вырезается поврежденный участок ДНК. Сначала фермент эндонуклеаза разрезает одну цепь. Следующий фермент — экзонуклеаза — удаляет измененную часть, Рис. 160. Механизм темновой репарации (этапы вырезания и репарации поврежденного участка молекулы ДНК): 1 — пиримидиновый димер искажает структуру ДНК, 2 — репарационная эндонуклеаза разрезает поврежденную цепь, 3 — репарационная эксцизия при действии экзонуклеазы, 4 — репарационный синтез, 5 — ДНК-лигаза заменяет оставшийся од-ноцепочечный разрыв.
а ДНК-полимераза вновь синтезирует недостающий участок, комплементарный неповрежденной цепи ДНК. И наконец, свободные концы старой части цепи соединяются с концами вновь синтезированного участка при помощи лигазы. Фотореактивация и эксцизионная репарация коротких участков ДНК протекают без ошибок и, следовательно, не ведут к мутациям. Если димеры не будут устранены, то соответствующие основания не смогут выполнять роль матрицы и в этих местах во вновь синтезированной цепи ДНК окажутся пропуски (пробелы, или бреши). Путем рекомбинации между двумя двойными цепями ДНК — продуктами репликации — возможно образование одной нормальной двойной цепи (пострепликативная репарация). Если повреждения лежат так тесно друг возле друга, что пропуски перекрываются, то для их заполнения используется другая «ремонтная» система (SOS-репарация), способная синтезировать новую цепь ДНК и на дефектной матрице. При этом новые основания включаются таким образом, что могут возникнуть мутации. Таким образом, репарации в значительной мере ограничивают количество мутаций, что имеет огромное биологическое значение. Однако не все виды повреждений ДНК репарируются, часть их проявляется в виде мутаций. В частности, известно несколько мутаций, проявляющихся как тяжелые наследственные заболевания за счет нарушения процесса репарации. Примером может служить пигментная ксеродерма — рецессивная аутосомная мутация — сверхчувствительность к ультрафиолетовым лучам под влиянием которых на открытых частях тела появляются пигментные пятна вначале в виде веснушек, затем все более крупных папиллом (родинок на ножках) различной величины и формы и, наконец опухоли.
|