Обмен веществами между клеткой и окружающей средой
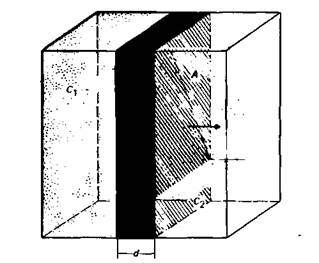
Мы кратко описали строение клетки, чтобы использовать это описание для рассмотрения основ клеточной физиологии. Клетку ни в коем случае нельзя считать статичным образованием, поскольку здесь происходит постоянный обмен веществами между различными внутриклеточными компартментами, а также между компартментами и окружающей средой. Структуры клетки находятся в динамическом равновесии, и взаимодействия клеток между собой и с внешней средой являются необходимым условием для поддержания жизни функционирующего организма. В данной главе мы рассмотрим фундаментальные механизмы такого обмена. В последующих главах эти механизмы будут рассмотрены в приложении к нервной клетке и ее функциям; однако те же самые механизмы лежат в основе функционирования всех других органов. Диффузия. Простейшим процессом перемещения вещества является диффузия. В растворах (или газах) атомы и молекулы перемещаются свободно, а разность концентраций уравновешивается благодаря диффузии. Рассмотрим два объема, заполненные жидкостью или газом (рис. 1.3), в которых вещества имеют концентрации с1 и с2 и разделены слоем с площадью поверхности А и толщиной d. Поток вещества m за время t описывается первым законом диффузии Фика.
где D - коэффициент диффузии, постоянный для данного вещества, растворителя и температуры. В более общем виде для разности концентраций dc на расстоянии dx
поток через сечение А пропорционален градиенту концентрации dc/dx. Знак «минус» возникает в уравнении потому, что изменение концентрации по направлению х - отрицательно. Диффузия - это наиболее важный процесс, благодаря которому б о льшая часть молекул в водных растворах перемещается на небольшие расстояния. Это относится и к их движению в клетке постольку, поскольку диффузии не препятствуют мембраны. Множество веществ могут свободно диффундировать через липидные мембраны, в особенности вода и растворенные газы, такие, как О2 и СО2. Жиро- 12 ЧАСТЬ I. ОБЩАЯ ФИЗИОЛОГИЯ КЛЕТКИ
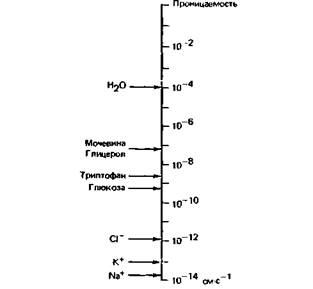
растворимые вещества также хорошо диффундируют через мембраны; это касается и полярных молекул достаточно малого размера, таких, как этанол и мочевина, тогда как сахара проходят через липидный слой с трудом. Вместе с тем липидные слои практически непроницаемы для заряженных молекул, в том числе даже для неорганических ионов. Для неэлектролитов уравнение диффузии (1) обычно преобразуют, соединяя характеристки мембраны и диффундирующего вещества в один параметр - проницаемость (Р):
На рис. 1.4 сравниваются проницаемости (Р) липидной мембраны для различных молекул. Диффузия через мембранные поры. Плазматическая мембрана (и другие клеточные мембраны) проницаемы не только для веществ, диффундирующих через липидный слой, но и для многих ионов, Сахаров, аминокислот и нуклеотидов. Эти вещества преодолевают мембрану через поры, образованные транспортными белками, погруженными в мембрану. Внутри таких белков имеется заполненный водой канал диаметром менее 1 нм, через который могут диффундировать малые молекулы. Они движутся по градиенту концентрации, и если они несут заряд, то их движение по каналам регулируется также мембранным потенциалом. Мембранные каналы обладают относительной избирательностью
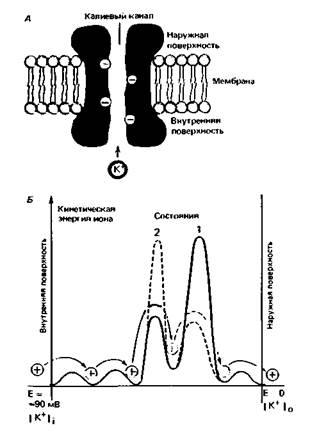
по отношению к типу молекул, которые могут через них проходить. Существуют, например, калиевые, натриевые и кальциевые каналы, каждый из которых непроницаем практически для любого иона, кроме специфического. Такая избирательность обусловлена зарядом или структурой мест связывания в стенках канала, что облегчает транспорт специфической молекулы и предотвращает проникновение через канал других веществ (рис. 1,5, А) [1, 3]. За поведением мембранных ионных каналов легко наблюдать, поскольку возникающий при движении ионов ток можно измерить, причем даже для одиночного канала. Показано, что каналы спонтанно и с высокой частотой меняют свое состояние от открытого к закрытому. Для калиевого канала характерны импульсы тока амплитудой около 2 пА (2-10 −12 А) и длительностью в несколько миллисекунд (см. рис. 2.12, стр. 37) [3]. За этот период через него проходят десятки тысяч ионов. Переход белков из одной конформации в другую изучают методами рентгеновской дифракции, мессбауэровской спектроскопии и ядерно-магнитного резонанса (ЯМР). Белки, таким образом, являются очень динамичными подвижными структурами, а канал, проходящий через белок, не просто жесткая, наполненная водой трубка (рис. 1.5, А), но лабиринт быстро двигающихся молекулярных групп и зарядов. Эта динамическая характеристика канала отражается в энергетическом профиле канала, показанном на рис. 1.5, Б. Здесь по оси абсцисс представлена длина канала от внешнего раствора с концентрацией ионов со и потенциалом 0 до внутреннего раствора с концентрацией с1 и потенциалом Е. По оси орди- ГЛАВА 1. ОСНОВЫ КЛЕТОЧНОЙ ФИЗИОЛОГИИ 13
нат представлены уровни энергии иона в местах связывания канала; пик на графике представляет барьер проницаемости, который энергия иона должна превосходить для проникновения через канал, а «провал» графика - сравнительно стабильное состояние (связывание). Несмотря на препятствие в виде энергетического пика, ион может проникнуть через канал, если энергетический профиль спонтан- но циклически меняется; ион, таким образом, может внезапно оказаться «по ту сторону» энергетического пика и может продолжить движение в клетку. В зависимости от заряда, размеров и степени гидратации иона и его способности связываться со структурами стенок канала энергетический профиль канала варьирует для различных ионов, чем может объясняться избирательность отдельных типов каналов. Диффузионное равновесие ионов. Диффузия различных ионов через мембранные каналы должна была бы привести к устранению различий в концентрациях между вне- и внутриклеточной средами. Как, однако, видно из табл. 1.1, такие различия сохраняются, следовательно, должно существовать некое равновесие между диффузией и другими процессами транспорта через мембрану. Следующие два раздела касаются тех путей, с помощью которых такое равновесие устанавливается. В случае ионов на диффузионное равновесие влияет их заряд. Диффузия незаряженных молекул обеспечивается разностью концентраций dc, и, когда концентрации уравниваются, собственно транспорт прекращается. На заряженные частицы дополнительно влияет электрическое поле. Например, когда ион калия выходит из клетки по градиенту своей концентрации, он несет один положительный заряд. Таким образом, внутриклеточная среда становится более отрицательно заряженной, в результате чего на мембране возникает разность потенциалов. Внутриклеточный отрицательный заряд препятствует выходу из клетки новых ионов калия, а те ионы, которые все же выйдут из клетки, еще больше увеличат заряд на мембране. Поток ионов калия прекращается, когда действие электрического поля компенсирует диффузионное давление вследствие разности концентраций. Ионы продолжают проходить через мембрану, но в равных количествах в обоих направлениях. Следовательно, для данной разности концентраций ионов на мембране существует равновесный потенциал Eion, при котором поток ионов через мембрану прекращается. Равновесный потенциал может быть легко определен с помощью уравнения Нернста:
(4) где R - газовая постоянная, Τ-абсолютная температура, z - валентность иона (отрицательная для анионов), a F - число Фарадея. Если подставить в уравнение константы, то при температуре тела (Т = 310К) равновесный потенциал для ионов калия Ек равен:
14 ЧАСТЬ I. ОБЩАЯ ФИЗИОЛОГИЯ КЛЕТКИ
Действительно, было обнаружено, что все клетки имеют мембранный потенциал; в мышечных клетках млекопитающих его уровень составляет около — 90 мВ. В зависимости от условий и относительных концентраций ионов клетки могут иметь мембранный потенциал в пределах от —40 до —120 мВ. Для клетки в приведенном выше примере (табл. 1.1) потенциал покоя, равный примерно —90 мВ, показывает, что потоки ионов калия через мембранные каналы находятся приблизительно в равновесии. Это неудивительно, поскольку в покоящейся мембране открытое состояние калиевых каналов наиболее вероятно, т.е. мембрана наиболее проницаема для ионов калия. Мембранный потенциал, однако, определяется потоками и других ионов. Легкость, с которой незаряженные частицы могут диффундировать через мембрану, количественно представлена в уравнении (3). Проницаемость для заряженных частиц описывается несколько более сложным уравнением:
где μ - подвижность иона в мембране, d - толщина мембраны, a R, Τ и F- известные термодинамические постоянные. Определенные таким образом значения проницаемости для различных ионов могут быть использованы для расчета мембранного потенциала Ет, когда ионы калия, натрия и хлора проходят через мембрану одновременно (с проницаемостью РK, PNa и РCl, соответственно). При этом предполагается, что потенциал падает в мембране равномерно, так что напряженность поля постоянна. В этом случае применяется уравнение Голдмана, или уравнение постоянного поля [6, 12]:
(7) Для большинства клеточных мембран РK приблизительно в 30 раз выше, чем PNa (см. также разд. 1.3). Относительная величина РС1 сильно варьирует; для многих мембран РС1 мала по сравнению с Рк, однако для других (например, в скелетных мышцах) РCl значительно выше, чем Рк. Активный транспорт, натриевый насос. В предыдущем разделе описаны пассивная диффузия ионов и образующийся вследствие этого мембранный потенциал при заданных внутри- и внеклеточных концентрациях ионов. Однако в результате этого процесса концентрация ионов внутри клетки стабилизируется не автоматически, поскольку мембранный потенциал несколько более электроотрицателен, чем Ек, и намного - по сравнению с ENa (около +60 мВ). Благодаря диффузии внутриклеточные концентрации ионов, по крайней мере калия и натрия, должны уравниваться с внеклеточными. Стабильность градиента ионов достигается посредством активного транспорта: мембранные белки переносят ионы через мембрану против электрического и (или) концентрационного градиентов, потребляя для этого метаболическую энергию. Наиболее важный процесс активного транспорта - это работа Na/K-насоса, существующего практически во всех клетках; насос выкачивает ионы натрия из клетки, одновременно накачивая ионы калия внутрь клетки. Таким образом обеспечивается низкая внутриклеточная концентрация ионов натрия и высокая - калия (табл. 1.1). Градиент концентрации ионов натрия на мембране имеет специфические функции, связанные с передачей информации в виде электрических импульсов (см. разд. 2.2), а также с поддержанием других активных транспортных механизмов и регулирования объема клетки (см. ниже). Поэтому неудивительно, что более 1/3 энергии, потребляемой клеткой, расходуется на Na/К-насос, а в некоторых наиболее активных клетках на его работу расходуется до 70% энергии [1, 11]. Na/К-транспортный белок представляет собой АТФазу. На внутренней поверхности мембраны она расщепляет АТФ на АДФ и фосфат (рис. 1.6). На транспортировку трех ионов натрия из клетки и одновременно двух ионов калия в клетку используется энергия одной молекулы АТФ, т. е. суммарно за один цикл из клетки удаляется один положительный заряд. Таким образом, Na/К-насос является электрогеиным (создает электрический ток через мембрану), что приводит к увеличению электроотрицательности мембранного потенциала приблизительно на 10 мВ. Транспортный белок выполняет эту операцию с высокой скоростью: от 150 до 600 ионов натрия в секунду. Аминокислотная последовательность транспортного белка известна, однако еще не ясен механизм этого сложного обменного транспорта. Данный процесс описывают с использованием энергетических профилей переноса белками ионов натрия или калия (рис. 1.5, Б). По характеру изменения этих профилей, связанных с постоянными изменениями конформации транспортного белка (процесс, требующий затраты энергии), можно судить о стехиометрии обмена: два иона калия обмениваются на три иона натрия. Na/К-насос, как и изолированная Na+/К+-зависимая мембранная АТФаза, специфически ингибируется сердечным гликозидом уабаином (строфантином). Поскольку работа Na/К-насоса представляет собой многоступенчатую химическую реакцию, она, подобно всем химическим реакциям, в значительной степени зависит от температуры, что проде- ГЛАВА 1. ОСНОВЫ КЛЕТОЧНОЙ ФИЗИОЛОГИИ 15
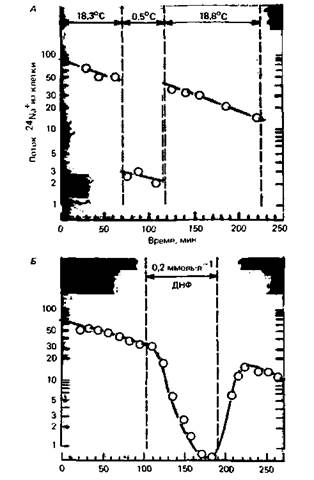
монстрировано на рис. 1.7. Здесь поток ионов натрия из мышечных клеток показан относительно времени; практически это эквивалентно потоку ионов натрия, опосредованному работой Na/K-насоса, потому что пассивный поток ионов натрия против градиентов концентрации и потенциала крайне мал. Если препарат охладить примерно на 18 oС, то поток ионов натрия из клетки быстро уменьшится в 15 раз, а сразу после нагревания восстановится до исходного уровня. Такое уменьшение потока ионов натрия из клетки в несколько раз больше, чем то, которое бы соответствовало температурной зависимости процесса диффузии или простой химической реакции. Сходный эффект наблюдается, когда запас метаболической энергии истощается в результате отравления динитрофенолом (ДНФ) (рис. 1.7, Б). Следовательно, поток ионов натрия из клетки обеспечивается энергозависимой реакцией-активным насосом. Другой характеристикой насоса наряду со значительной температурной и энергетической зависимостью является наличие уровня насыщения (как и у всех других химических реакций): это означает, что скорость работы насоса не может возрастать бесконечно при повышении концентрации транспортируемых ионов (рис. 1.8). В отличие от этого поток пассивно диффундирующего вещества растет пропорционально разности концентраций в соответствии с законом диффузии (уравнения 1 и 2).
Помимо Na/К-насоса плазматическая мембрана содержит по крайней мере еще один насос -кальциевый; это насос откачивает ионы кальция (Са2+) из клетки и участвует в поддержании их внутриклеточной концентрации на крайне низком уровне (табл. 1.1). Кальциевый насос присутствует с очень высокой плотностью в саркоплазматическом ретикулуме мышечных клеток, которые накапливают ионы кальция в результате расщепления молекул АТФ (см. гл. 4). Воздействие Na/K-насоса на мембранный потенциал и объем клетки. На рис. 1.9 показаны различные компоненты мембранного тока и приведены внутриклеточные концентрации ионов, которые обе- 16 ЧАСТЬ I ОБЩАЯ ФИЗИОЛОГИЯ КЛЕТКИ
спечивают их существование. Через калиевые каналы наблюдается выходящий ток ионов калия, так как мембранный потенциал несколько более электроположителен, чем равновесный потенциал для ионов калия. Общая проводимость натриевых каналов намного ниже, чем калиевых, т. е. натриевые каналы открыты намного реже, чем калиевые при потенциале покоя; однако в клетку входит примерно столько же ионов натрия, сколько выходит из нее ионов калия, потому что для диффузии ионов натрия в клетку необходимы большие градиенты концентрации и потенциала. Na/K-насос обеспечивает идеальную компенсацию пассивных диффузионных токов, так как переносит ионы натрия из клетки, а ионы калия - в нее. Таким образом, насос является электрогенным за счет разницы в числе перенесенных в клетку и из клетки зарядов, что при нормальной скорости его работы создает мембранный потенциал, примерно на 10 мВ более электроотрицательный, чем если бы он образовывался только за счет пассивных потоков ионов (см. уравнение 7). В результате мембранный потенциал приближается к калиевому равновесному потенциалу, что уменьшает утечку ионов калия. Активность Na/K-насоса регулируется внутриклеточной концентрацией ионов натрия. Скорость работы насоса замедляется при снижении концентрации ионов натрия, подлежащих выводу из клетки (рис. 1.8), так что работа насоса и поток ионов натрия внутрь клетки уравновешивают друг друга, поддерживая внутриклеточную концентрацию ионов натрия на уровне примерно 10 ммоль-л—1. Чтобы поддерживать равновесие между насосными и пассивными мембранными токами, необходимо намного больше молекул Na/K-насоса, чем канальных белков для ионов калия и натрия. При открытом состоянии канала через него проходят десятки тысяч ионов за несколько миллисекунд (см. выше), а поскольку канал обычно открывается несколько раз в секунду, всего за это время через него проходит более 105 ионов. Одиночный насосный белок перемещает несколько сотен ионов натрия в секунду, следовательно, плазматическая мембрана должна содержать примерно в 1000 раз больше насосных молекул, чем канальных. Измерения канальных токов в покое показали наличие в среднем одного калиевого и одного натриевого открытого канала на 1 мкм2 мембраны; из этого следует, что на том же пространстве должно присутствовать около 1000 молекул Na/K-насоса, т.е. расстояние между ними составляет в среднем 34 нм; диаметр насосного белка, как н канального, составляет 8-10 нм. Таким образом, мембрана достаточно плотно насыщена насосными молекулами [11]. Тот факт, что поток ионов натрия внутрь клетки, а ионов калия - из клетки компенсируется работой насоса, имеет и другое следствие, заключающееся в сохранении стабильного осмотического давления и постоянного объема. Внутри клетки существует высокая концентрация крупных анионов, главным образом белков (А— в табл. 1.1), которые не способны проникать через мембрану (или проникают через нее очень медленно) и поэтому являются фиксированным компонентом внутри клетки. Чтобы уравновесить заряд этих анионов, необходимо равное количество катионов. Благодаря действию Na/K-насоса этими катионами в основном оказываются ионы калия. Существенное повышение внутриклеточной концентрации ионов могло бы происходить только при возрастании концентрации анионов вследствие потока С1— по градиенту концентрации в клетку (табл. 1.1), но мембранный потенциал противодействует этому. Входящий ток С1— наблюдается только до тех пор, пока не будет достигнут равновесный потенциал для ионов хлора; ГЛАВА 1. ОСНОВЫ КЛЕТОЧНОЙ ФИЗИОЛОГИИ 17 это наблюдается, когда градиент ионов хлора практически противоположен градиенту ионов калия, так как ионы хлора заряжены отрицательно (уравнение 4). Таким образом, устанавливается низкая внутриклеточная концентрация ионов хлора, соответствующая низкой внеклеточной концентрации ионов калия. Результатом является ограничение общего количества ионов в клетке. Если мембранный потенциал падает при блокаде Na/К-насоса, например при аноксии, то равновесный потенциал для ионов хлора снижается, а внутриклеточная концентрация ионов хлора соответственно повышается. Восстанавливая равновесие зарядов, ионы калия также входят в клетку; суммарная концентрация ионов в клетке возрастает, что повышает осмотическое давление; это заставляет воду поступать в клетку. Клетка набухает. Такое набухание наблюдается in vivo в условиях недостатка энергии. Концентрационный градиент Na+ как движущая сила мембранного транспорта. Значение Na/К-насоса для клетки не ограничивается стабилизацией нормальных градиентов К+ и Na+ на мембране. Энергия, запасенная в мембранном градиенте Na+, часто используется для обеспечения мембранного транспорта других веществ. Например, на рис. 1.10 показан «симпорт» Na+ и молекулы сахара в клетку. Мембранный транспортный белок переносит молекулу сахара в клетку даже против градиента концентрации, в то же время Na+ движется по градиенту концентрации и потенциала, обеспечивая энергию для транспорта Сахаров. Такой транспорт Сахаров полностью зависит от существования высокого градиента Na+; если внутриклеточная концентрация Na+ существенно возрастает, то транспорт Сахаров прекращается. Для различных Сахаров существуют разные симпортные системы. Транспорт аминокислот в клетку сходен с транспортом Сахаров, показанным на рис. 1.10; он также обеспечивается градиентом Na+; существует по крайней мере пять различных систем" симпорта, каждая из которых специализирована для какой-либо одной группы родственных аминокислот. Помимо симпортных систем существуют также «антипортные». Одна из них, например, за один цикл переносит один ион кальция из клетки в обмен на три входящих иона натрия (рис. 1.10). Энергия для транспорта Са2+ образуется за счет входа трех ионов натрия по градиенту концентрации и потенциала. Этой энергии достаточно (при потенциале покоя) для поддержания высокого градиента ионов кальция (от менее 10 −7 моль-л−1 внутри клетки до приблизительно 2 ммоль-л−1 вне клетки). Эндо- и экзоцитоз. Для некоторых веществ, которые поступают в клетку или должны быть выведены
из нее, транспортные каналы отсутствуют; к таким веществам относятся, например, белки и холестерол. Они могут проходить через плазматическую мембрану в везикулах,или пузырьках,с помощью эндо- и экзоцитоза. На рис. 1.11 показаны основные механизмы этих процессов. При экзоцитозе определенные органеллы (см. ниже) формируют везикулы, заполненные веществом, которое необходимо вывести из клетки, например гормонами или ферментами внеклеточного действия. Когда такие везикулы достигают плазматической мембраны, их липидная мембрана сливается с ней, давая таким образом возможность содержимому выйти во внешнюю среду. При противоположном процессе - эндоцитозе - плазматическая мембрана инвагинирует, образуя ямку, которая затем углубляется и замыкается, формируя внутриклеточную везикулу, заполненную внеклеточной жидкостью и некоторыми макромолекулами. Чтобы обеспечить это слияние мембран и замыкание везикулы, сократительные элементы цитоскелета действуют совместно с самими мембранами (см. ниже). При эндоцитозе не всегда происходит просто захват внеклеточной среды в клетку. В клеточной мембране содержатся часто организованные в специализированные группы специфические рецепторы к макромолекулам, таким, как инсулин или антигены. После того как эти макромолекулы свяжутся со своими рецепторами, в окружающем рецептор участке мембраны происходит эндоцитоз, и макромолекула избирательно транспортируется в клетку (рис. 1.12, Б). Эндо- и экзоцитоз происходят в клетках непрерывно. Количество мембранного материала, совершающего оборот, значительно; в течение 1 ч макрофаг поглощает в виде везикул двойную площадь поверхности своей цитоплазматической мембраны. В большинстве клеток оборот мембранного материала происходит не столь интенсивно, но все же должен быть значительным. 18 ЧАСТЬ I. ОБЩАЯ ФИЗИОЛОГИЯ КЛЕТКИ
|
 (1)
(1) (2)
(2)
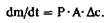 (3)
(3)
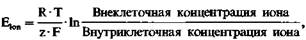
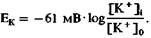 (5)
(5)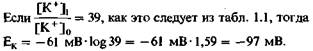
 (6)
(6)