
Формы транспорта СО2
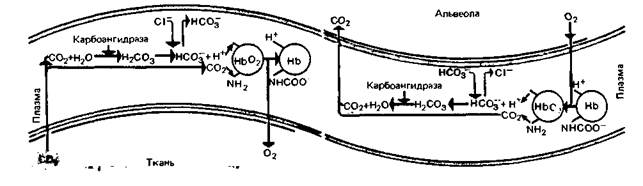
Диоксид углерода (СО2, углекислый газ)-конечный продукт окислительного метаболизма в клетках - переносится с кровью к легким и удаляется через них во внешнюю среду. Подобно кислороду, диоксид углерода может переноситься как в физически растворенном виде, так и в составе химических соединений. Химическое связывание СО2 - более сложный процесс по сравнению со связыванием кислорода. Это обусловлено тем, что механизм, отвечающий за транспорт СО2, должен одновременно обеспечивать поддержание постоянства кислотно-щелочного равновесия крови и тем самым внутренней среды организма в целом. Связывание СО2 [15]. Напряжение СО2 в артериальной крови, поступающей в тканевые капилляры, составляет 40 мм рт. ст. (5,3 кПа). В клетках же, расположенных около этих капилляров, напряжение СО2 значительно выше, так как углекислый газ постоянно образуется в процессе метаболизма. В связи с этим физически растворенный СО2 диффундирует по градиенту напряжения из тканей в капилляры. Здесь некоторое количество углекислого газа остается в растворенном состоянии, но большая часть СО2 претерпевает ряд химических превращений (рис. 22.9). Прежде всего происходит гидратация молекул СО2 с образованием угольной кислоты, сразу же диссоциирующей на ион бикарбоната и протон:
В плазме крови эта реакция протекает очень медленно; в эритроците же она ускорена примерно в 10 тыс. раз. Это связано с действием фермента карбоангидразы [6, 24]. Поскольку этот фермент ГЛАВА 22. ТРАНСПОРТ ГАЗОВ КРОВИ 615
присутствует только в эритроцитах, практически все молекулы СО2, участвующие в реакции гидратации, должны сначала проникнуть в эритроциты. Накопление в эритроците НСО3− приводит к тому, что между его внутренней средой и плазмой крови создается диффузионный градиент. Ионы НСО3− могут двигаться по этому градиенту лишь в том случае, если при этом не нарушается равновесное распределение электрических зарядов. Поэтому одновременно с выходом каждого иона НСО3− должен происходить либо выход из эритроцита одного катиона, либо вход одного аниона. Поскольку мембрана эритроцита практически непроницаема для катионов, но сравнительно легко пропускает небольшие анионы, в обмен на НСО3− в эритроцит поступают ионы С1~. Этот обменный процесс называют хлоридным сдвигом (сдвигом Хамбургера). По мере поступления СО2 в эритроците образуются не только ионы НСО3−, но также ионы Η +. Однако это не приводит к значительным сдвигам pH внутри эритроцита, что обусловлено, в частности, особыми свойствами гемоглобина. Это вещество, будучи амфолитом, обладает значительной буферной емкостью. Кроме того, восстановленный гемоглобин обладает более слабыми кислотными свойствами, чем оксигемоглобин, поэтому он может присоединять дополнительное количество ионов Η + (с. 619). СО2 может связываться также путем непосредственного присоединения к аминогруппам белкового компонента гемоглобина. При этом образуется остаток карбаминовой кислоты (карбамат):
Гемоглобин, связанный с СО2, называется карбаминогемоглобином (или упрощенно карбогемоглобином). Все эти химические реакции и взаимосвязи между ними представлены на рис. 22.9. В левой части этого рисунка показаны процессы, происходящие в тканевых капиллярах при поступлении СО2 в кровь. В правой части изображены реакции, протекающие при высвобождении СО2 в легких; видно, что направление всех реакций здесь противоположное. Роль разных форм СО2 в газообмене. В крови, поступающей к тканям, напряжение СО2 составляет 40 ммрт. ст. Проходя через них, кровь насыщается углекислым газом, и напряжение его в оттекающей из тканей крови достигает в среднем 46 мм рт. ст. При этом 1 л крови поглощает примерно 1.8 ммоль СО2. Около 12% этого количества остается в физически растворенном виде или в форме недиссоциированной угольной кислоты (Н2СО3), 11% образует карбаминовое соединение с гемоглобином, 27% транспортируется в виде бикарбоната в эритроцитах, а остальное количество-около 50%-растворено в виде НСО3− в плазме. При прохождении крови через легкие СО2 высвобождается из этих четырех форм в таком же соотношении. Сатурационные кривые CO2 1)
|
 (9)
(9)
 (10)
(10)


