
Вторинно - активне транспортування
Найчастіше цей вид перенесення речовин забезпечується у поєднанні з транспортуванням Nа+ (наприклад, транспорт вуглеводів і амінокислот ентероцитами тонкого кишечника, іонів Са2+ в кардіоміоцитах). Потенціал спокою, його параметри, механізми походження, фізіологічна роль Потенціал спокою (МПС) – це різниця потенціалів між зовнішньою та внутрішньою поверхнями мембрани збудливої клітини, що знаходиться в стані спокою. Теорій виникнення МПС багато (Хожкін, Хакслі, Берштейн), але суть одна - підґрунтям для виникнення МПС є неоднакова проникність мембрани для катіонів і аніонів, внаслідок чого виникає асиметрія розподілу іонів в клітині між зовнішньою і внутрішньою поверхнею мембрани. Зовнішня поверхня має «+» заряд, а внутрішня «-».
Механізм формування МПС пов'язаний з: 1. Наявністю в мембрані клітини механізмів активного транспорту речовин – натрій-калієвого насосу. Він створює градієнт концентрації усередині та поза клітиною. 2. Особливостями проникності мембрани клітини в стані спокою такі: вона проникна для іонів К+ та непроникна для іонів натрію.
Структурною одиницею НКН є транспортний білок, що являє собою АТФазу. Цей білок на внутрішній поверхні мембрани розщеплює АТФ на АДФ та фосфат й використовує енергію, що виділилась на транспортування трьох іонів натрію з клітини та двох іонів калію в клітину. Тобто в сумі з клітини за один цикл виділяється один позитивний заряд (150-600 за секунду). Таким чином, НКН є електрогенним (створює електричний струм через мембрану), але може бути електронейтральним. Проникність мембрани для іонів визначається станом каналів мембрани. В стані спокою мембрана збудливої клітини проникна для іонів К+ (відкриті калієві канали) та непроникна для іонів Nа+ - майже всі натрієві канали закриті. Це є причиною того, що іони К+ виходять з клітини через калієві канали по градієнту концентрації, а іони Na+ зайти в клітину не можуть, хоча є градієнт концентрації. Іони К+, виходячи з клітини, виносять позитивний заряд на зовнішню поверхню мембрани. Крупномолекулярні органічні аніони залишаються в клітині й зумовлюють негативний заряд на внутрішній поверхні мембрани.
Величина МПС в стані спокою різна, але постійна в нормі: - скелетний мяз МПС- 90мВ; - гладенький мяз - ЗОмВ; - нервова клітина - 40 - 90мВ; - секреторна клітина - 20мВ. Методи реєстраціїМПС Для вивчення електричних властивостей клітинних мембран використовують різні прилади і методики.
При реєстрації ПС методом пошкодження об'єктом дослідження є який-небудь орган, наприклад, скелетний м'яз. Його пошкоджують шляхом розрізу і прикладають електроди до пошкодженої та непошкодженої поверхонь. Стрілка гальванометра при цьому відхиляється. Це говорить про наявність різниці потенціалів між пошкодженою поверхнею м'яза (аналог внутрішньоклітинного середовища) та її непошкодженою поверхонею (аналог зовнішньої мембрани).
При внутрішньоклітинному методі реєстрації об'єктом дослідження є окрема клітина, яку поміщають в плазму чи фізіологічний розчин. Там знаходиться один реєструючий електрод. Другий електрод обережно вводиться усередину клітини. До проколу мембрани промінь осцилографа іде по нульовій лінії бо між електродами немає різниці потенціалів. В момент проколу мембрани промінь приладу різко відхиляється від 0 у негативну частину системи координат і протягом годин може перебувати на такому рівні. Фізіологічна рольМ ПС полягає в тому, що він є базою, на якій розвивається потенціал дії, тобто МПС забезпечує біологічну властивість - збудливість, тобто готовність до відповіді на подразнення.
Потенціл дії, його фази, механізм утворення, методи реєстрації, параметри ПД
В основі зміни мембранного потенціалу при збуджені (тобто в основі формуванні ПД) лежать зміни проникності мембрани, що пов'язані із змінами стану каналів мембрани під впливом подразника. Подразник зумовлює відкриття активаційних воріт натрієвих каналів і відбувається лавиноподібний вхід Na + в клітину за градієнтом концентрації, це призводить до зменшення надлишку аніонів усередині клітини Аніони, які прямують до мембрани за іонами Na, зайти в клітину не можуть. Завдяки цьому різниця потенціалів між зовнішньою та внутрішньою поверхнями мембрани зменшується, а далі й зовсім зникає до 0.Так розвивається фаза деполяризації. Але вхід іонів Na + в клітину не припиняється тому у клітині створюється надлишок позитивних іонів Na +, а на її поверхні - надлишок аніонів відбувається перезарядка
мембрани - фаза реверсполяризації. Натрієва активація (стан каналів при відкритих активаційних воротах) швидко змінюється натрієвою інактивацією - закриттям інактиваційних воріт, вхід іонів Na + в клітину при цьому припиняється. Подразник змінює не тільки стан натрієвих каналів, але й стан калієвих каналів, вони також активуються, тобто відкривається додаткова кількість калієвих каналів Тому, коли іони Na + перестають входити в клітину, вихід іонів К з неї збільшується (пасивно). Саме це призводить до накопичення «+» зарядів на мембрані, відновлення вихідного МПС, ця фаза називається реполяризація. Таким чином, механізм формування ПД полягає в наступному: -відкриття і закриття Na+, К+ каналів під час розвитку ПД відбувається під впливом МП, тому ці канали називають потенціалзалежними. -ПД, порушує іонні градієнти, властиві для спокійної клітини, тому для відновлення іонних градієнтів у фазі реполяризації активується Na - К+ насос, який відновлює іонний склад клітини до вихідного рівня. Таким чином, виникнення ПД, відбувається внаслідок пасивного проходження іонів через мембрану, а відновлення іонних градієнтів Na+, К+ потребує енергії (АТФ), роботи натрій- калієвого насоса. Спочатку Na+ канал викачує Na+ назовні, а К + канал повертає К + в середину клітини. Це завершує відновлення МПС (- 90 мВ). Дія К +каналу продовжується деякий час, тому К+ входить в клітину більше, ніж вийшло, тому виникає різниця між внутрішньою і зовнішньою поверхнями мембрани - 93 - 95 мВ проти 90 мВ - це гіперполяризація, є позитивною частиною слідового потенціалу триває 50 - 300 мс. Параметри ПД: 1.Амплітуда- 100-120 мВ. 2.Протяжність - 1-3 мс в нервових волокнах, в м'язових волокнах він довший (в типових кардіоміоцитах - 300 мс Фізіологічна роль ПД - полягає в тому, що він забезпечує збудження клітини та передачу цього збудження на інші клітини, а також підтримку чи активацію у збудливих клітинах специфічних функцій. Механізм проведення нервового імпульсу по нервовим волокнам (мієліновим та безмієліновим).Швидкість проведення збудження
Нервові волокна -це відростки нейронів. Нервове волокно складається із осьового циліндра, покритого оболонками. За будовою всі нервові волокна поділяються на: - безмієлінові, вони не мають мієлінової оболонки, тому збудження виникає в любому місці волокна, швидкість проведення імпульсу низька 0, 5 - 2 м/сек - мієлінові волокна, вкриті мієліновою оболонкою, яка складається із жироподібної речовини і є електричним ізолятором. Через певні проміжки (0, 2 - 2 мкм) мієлінова оболонка переривається ділянками, шириною приблизно 1 мкм, - це перехвати Ранв'є.Перехвати Ранв'є пронизані багаточисельними каналами, тому в цих, оголених від мієліну місцях, виникає ПД.Швидкість проведення імпульсів висока 80-120 м/сек.
Механізм проведення збудження по безмієліновим волокнам В теперішній час теорія кругових струмів підтверджена експериментально: між збудженою і не збудженою ділянками струм рухається від «+» до «-», це призводить до малих колових струмів, які захоплюють нові ділянки, у безмієлінових волокнах збудження поширються безупинно уздовж мембрани осьового циліндра.
.
Під впливом подразника (П) на мембрані волокна виникає збудження - ПД. Деполяризація мембрани йде послідовно, захоплює все нові ділянки і веде до того, що ПД поширюється без зміни амплітуди зі швидкістю 0, 5 - 2 м/с. Параметри цих струмів як подразника: 1. Напрям - з рисунка видно, що струми мають вихідний (катодний) напрям в незбуджених ділянках мембрани. Отже, тут буде виникати деполяризація мембрани. Якщо вона дійде до Екр, то виникне ПД. 2. Сила - чисельно сила струму в даному випадку рівна різниці потенціалів між збудженими та незбудженими ділянками мембрани і ця сила відповідає амплітуді ПД 3. Час дії подразника - відповідає тривалості ПД. 4. Швидкість збільшення сили - відповідає швидкості піку ПД. Проведення збудження по мієліновим нервовим волокнам Механізм проведення збудження по мієліновим нервовим волокнам принципово не відрізняється від механізму проведення по безмієлінових волокнах: під впливом подразника (П) в одному з перехватів Ранв'є виникає ПД.Між цим (збудженим) і сусідніми(незбудженими) перехватами Ранв'є виникає різниця потенціалів дії; вони з'єднані провідним середовищем, тому виникають місцеві струми (від " +" до " -").
Суттєва відмінність поширення ПД по мієліновим волокнам полягає в тому, що в них місцеві струми виникають не між сусідніми ділянками мембрани, а між сусідніми перехватами Ранв'є, тому ПД поширюється від одного перехвату до іншого стрибкоподібно і швидкість поширення ПД збільшується. При сальтаторному проведенні збудження швидкість дорівнює 80-120 м/сек. На швидкість поширення збудження по нервовим волокнам впливають такі фактори: 1. Наявність мієлінової оболонки збільшують швидкість. 2. Відстань між перехватами Ранв'є, чим вона більша, тим більша швидкість. 3. Діаметр волокна - чим він більший, тим менший опір чинить аксоплазма волокна поширенню локальних струмів і тим більша швидкість проведення збудження. 4. Амплітуда ПД - чим вона більша, тим швидше деполяризація доходить до Екр, і тим більша швидкість проведення. 5. Поріг деполяризації - чим він менший, тим виникає швидше деполяризація мембрани. 6. Швидкість наростання піку ПД - чим вона більша, тим швидше розвивається деполяризація до Екр, і тим більша швидкість проведення збудження. Закони проведення збудження по нервовим та м 'язовим волокнам 1. Закон фізіологічної неперервності чи фізіологічної цілісності волокна - для здійснення проведення необхідним є нормальний функціональний стан мембрани волокна. Якщо волокно пошкодити, обробивши, наприклад, місцевим анастетиком типу новокаїну, проведення збудження припиниться, незважаючи на те, що морфологічно волокна не пошкоджені (місцеві анастетики блокують натрієві канали мембрани). 2. Закон двобічного проведення - в умовах експерименту збудження (ПД) поширюється вздовж волокна в дві сторони, тобто, провідність волокон двобічна. Але в умовах цілісного організму відбувається одностороннє проведення збудження по нервовим волокнам, незважаючи на двосторонню провідність - ПД поширюється від рецепторів до нервових центрів, від нервових центрів до органів-ефекторів. Це відбувається завдяки тому, що обов'язковим елементом будь-якої рефлекторної дуги є синапси, які мають односторонню провідність. 3. Закон ізольованого проведення - ПД, поширюючись по мембрані одного волокна, не передається на сусідні й не викликає ПД на мембрані сусідніх волокон. Приклад, сідничний нерв має тисячі волокон, якби не було ізольованого проведення імпульсу, то виникає хаотична відповідна реакція. 4. Закон бездекрементного проведення - амплітуда ПД не змінюється при проведенні збудження по мембрані волокна. Причиною цього є те, що ПД підкоряється закону " все або нічого" - в кожній ділянці мембрани він має максимально можливу для цього волокна амплітуду. 5. Проведення без розвитку втоми. Нервові волокна практично не стомлюються - на протязі багатьох годин відтворюють частоту стимуляції.. Введенський довів, що нерв можна подразнювати безперервно 9-12 год. 6.Проведення з дуже високим коефіцієнтом надійності.
Механізми утворення та передачі збудження, Властивості і будова м м’язового волокна Посмуговані м'язи складається із м'язових волокон - це тонкі(< 3-0, 01-0, 1мм), витягнуті (2-Зсм, до 10см), багатоядерні утворення в результаті злиття багатьох клітин - симпласт. Ядра знаходяться біля поверхні волокна, ядер близько 1 тис. Оболонка називається сарколема, а цитоплазма - саркоплазма. В саркоплазмі волокна знаходиться велика кількість, приблизно 2500, міофібрилів. Саркоплазма волокна має мережу внутрішніх мембран - це саркоплазматичний ретикулюм, він складається з трубочок і пухирців(цистерн), які є депо кальція, на мембранах СР працює Са2+ насос. Міофібрили знаходяться у волокні в великій кількості і мають ділянки темні і світлі, їх називають диски, в зв'язку з цим волокно має поперечну смугастість. Між міофібрилами є багато мітохондрій. Міофібрила (міофіламент) складається із протофібрил, тобто білкових ниток 2 – х типів: -актинові, тонкі, 4, 5 – 6, 5 мкм., вони мають білок тропінін, кальмодулін, що закривають активний центр актину, але під дією Са2+ він відкривається, тому можуть утворюватись актиноміозинові містки; -міозинові, товсті, 10 – 15 мкм, мають хвіст, шийку і голівку, яка приймає участь в утворенні актиноміозинових містків. Під світловим мікроскопом міофібрили мають темні і світлі смуги - диски (А і І) - це пояснюється особливим розміщенням актинових і міозинових ниток.
Саркомер - це ділянка міофібрил розташованих між двома Z пластинами, довжина приблизно 25кмк.
Спряження збудження та скорочення. Механізм м’язового скорочення Термін " спряження збудження із скороченням" означає взаєзв'язок збудження в скелетних м'язах (виникнення та поширення ПД по мембрані волокна) та його скорочення
В стані спокою взаємодії актинових та міозинових протофібрил немає тому, що не знімають блокаду іони Са2+, концентрація яких в саркоплазмі в стані спокою низька 10~ ммоль/л. ПД, що виникають в постсинаптичній мембрані поширюються вздовж всієї довжини мембрани м'язового волокна, у томучислі і по мембрані СПР.При русі ПД по мембрані Т-трубочок, цистерн СПР відкриваються кальцієві канали, іони Са2 по градієнту концентрації виходять з цистерн СПР у саркоплазму, в результаті чого відбувається підвищення концентрації іонів Са2+ в саркоплазмі міозитів. Іони Са2+ взаємодіють з білком тропоніном, в результаті чого звільняються активні центри і утворюються актиноміозинові містки, тобто головка міозину з’єднується з активним центром актину. В основі м'язового скорочення лежить ковзання актинових протофібрил відносно міози нових, " теорія ковзання".
Для розслаблення скелетного м'язу необхідно, щоб зменшилась концентрація іонів Са2+ в саркоплазмі. Це досягається завдяки дії кальцієвих насосів повздовжніх трубочок СПР, які перекачують туди Са2+. Під час розслаблення м’язу відбувається: - активний транспорт іонів Са2+ в СПР; - зменшення його концентрації в саркоплазмі; - від'єднання іонів Са2+ від регуляторного білка тропініну, - актиміозинові зв’язки розиваються; - відбувається відновлення вихідної структури і відновлення положення тропоніну та тропоміозину, шо призводить до блокади активних центрів актинових ниток.
Будова і функції нервово – м’язового синапса
Будова нервово-мязового синапаса: -Нервовое волокно, яке підходить до м’язової клітини (волокна), втрачає мієлін і розширюється – це нервове закінчення або синаптична бляшка. -Мембрана нервового закінчення, яка прилягає до м’язового волокна називається пресинаптичною мембраною. -Постсинаптична мембрана, або кінцева пластинка, це мембрана м’язового волокна, що прилягає до нервового закінчення. -Пресинаптична і постсинаптична мембрани розділені синаптичною щілиною. -Нервове закінчення має синаптичні везикули заповнені АХ, Са2+ в нервовому закінченні менше, ніж у міжклітинній рідині. -На постсинаптичній мембрані розташовані хемозбудливі натрієві канали, які активуються АХ, що зв’язується з хеморецепторами.
полягає в тому, що ПД іде по мембрані нервового волокна і поширюється по пресинаптичній мембрані, при цьому відкриваються кальцієві канали пресинаптичної мембрани і відбувається вхід іонів Са всередину нервового закінчення та взаємодія з везикулами, які рухаються до пресинаптичної мембрани, відбувається злиття везикул з пресинаптичною мембраною, тому медіатор ацетилхолін входить в синаптичну щілину порціями. Дифузія ацетилхоліну до мембрани кінцевої пластинки призводить до взаємодії АХ з мембранними циторецепторами (Н-холінорецептори) і відкриття хемочутливих натрієвих каналів, тому іони Na+ пасивно входять в м'язове волокно через кінцеву пластинку, що викликає розвиток деполяризації кінцевої пластинки, що має назву потенціалу кінцевої пластинки (ПКП). ПКП або збудливий постсинаптичний потенціал (ЗПСП) має властивості локальної відповіді: не поширюється на мембрані; не підлягає закону «все або нічого».
Чим більше виділяється медіатора, тим більше ЗПСП. У разі досягнення критичного рівня деполяризації, струми, що виникають між деполяризованою постсинаптичною мембраною і сусідніми ділянками м’язового волокна, викликають генерацію ПД, який поширюється мембраною м’язового волокна і починається процес м’язового скорочення. Закономірності проведення збудження через нервово-м 'язовий синапс: 1.Однобічне проведення. 2. Уповільнене проведення (час від виникнення ПД на пресинаптичній мембрані до виникнення ПД на постсинаптичній мембрані складає близько 0, 5 мсек. 3. Проведення збудження через синапс супроводжується швидким розвитком втоми (у зв'язку з виснаженням запасів медіатора в нервовому закінченні); Ці особливості (1-3) характерні для будь-якого хімічного синапса і пов'язані з хімічним характером передачі інформації через синапс. 4.Збудження передається через нервово-м'язовий синапс без трансформації ритму. Типи м’язових скорочень: одинокі та тетанічні; ізотонічні та ізометричні
В залежності від режимів навантаження виділяють наступні види м'язового скорочення.: 1. Ізометричне скорочення – це скорочення при незмінній довжині м'яза.Довжина м'яза при скороченні залишається сталою, але зростає її напруження (Н). В експерименті таке скорочення можна отримати, якщо ізольований м'яз закріпити з двох сторін та стимулювати електричним струмом. В умовах цілісного організму ізометричне скорочення буває, коли людина намагається, але не може підняти вантаж. 2. Ізотонічне скорочення - скорочення при незмінному напруженні м'яза. Напруження м'яза при його скороченні залишається сталим, а довжина змінюється. В експерименті таке скорочення можна отримати, якщо один кінець ізольованого м'яза закріпити, а інший залишити вільним та стимулювати. В умовах цілісного організму скорочення м'язів максимально наближується до ізотонічного, якщо відбувається згинання ненавантажених кінцівок. 3. Змішане (ауксотонічне) скорочення - скорочення, при якому змінюється довжина та напруження м'яза. Такий тип м'язових скорочень найчастіше зустрічається в умовах цілісного організму. В залежності від частоти стимуляції виділяють наступні види м'язового скорочення:
1. Одиночне - спостерігається у відповідь на одиничний стимул чи на їх серію (якщо інтервал між подразниками більший чи рівний тривалості одиночного м'язового скорочення). Виділяють такі періоди м'язового скорочення:
- період скорочення (2) - під час цього періоду довжина м'яза зменшується; - період розслаблення (3) — під час цього періоду м'яз розслаблюється, його довжина збільшується до вихідного рівня. 2. Тетанічне - спостерігається в умовах сумації одиночних м'язових скорочень (одне скорочення накладається на інше). Така сумація можлива, якщо м'яз подразнюється серією стимулів і кожен наступний стимул діє тоді, коли попереднє одиночне скорочення ще не закінчилось (тобто, інтервал між стимулами має бути меншим тривалості одиночного м'язового скорочення). Види тетанічних скорочень: Зубчастий тетанус - виникає на базі одиночних скорочень, якщо кожен наступний стимул попадає на період розслаблення одиночого м'язового скорочення. Отже, інтервал між стимулами в серії має бути менший тривалості одиночного м'язового скорочення, але більший, чим сума тривалості латентного періоду і періоду вкорочення одиночного скорочення. Гладкий тетанус - виникає на базі одиночних скорочень, якщо кожен наступний стимул попадає на період скорочення.
Стрілочками показано момент дії подразника на м’язове волокно.
Асинхронний тетанус. Асинхронний тетанус представляє вид скорочень, який притаманний скелетним м'язам при їх природній діяльності в умовах цілісного організму. Кожен скелетний м'яз складається з великої кількості окремих м'язових волокон. При природній діяльності м'язів в умовах цілісного організму частота, з яким до них надходять ПД по аксонах мотонейронов недостатня для того, щоб у кожному окремому волокні м'яза розвивався гладкий тетанус. У той же час м'язи людини здатні до тривалого злитого скорочення. Таке скорочення є наслідком асинхронного (неодночасного) скорочення окремих м'язових волокон м'яза. Кожне волокно скорочується в режимі одиночних скорочень, але в межах м'яза відбувається (додавання) скорочень окремих волокон. У підсумку м'яз скорочується злитно та довготривало. Це і є асинхронний тетанус.
Рухові одиниці Рухове волокно (це аксон мотонейрона спинного мозку) у м’язі розгалужується і іннервує групу м’язових волокон. Ці м’язові волокна скорочуються одночасно під час надходження нервового імпульсу по аксону до м’яза. Мотонейрон + група м’язових волокон, які він іннервує називається рухова (моторна) одиниця. Розрізняють такі види рухових одииць: - швидкі РО; - повільні РО: вони скорочуються триваліший час, швидкість скорочення менша, м’язова сила менше. Втома розвивається пізніше. - Змішані тому, що мають швидкі і повільні РО. Але є м’язи, які складаються переважно: - із швидких РО, вони називаються швидкі, білі (прямий м’яз ока); - із повільних РО, вони називаються червоні, повільні (камбаловидний м’яз гомілки); М’язові волокна однієї моторної одиниці збуджуються одночасно, синхронно. М’язові волокна в цілому у м’язі працюють асинхронно, бо різні рухові одиниці іннервуються різними нейронами, які посилають імпульс з різною частотою. Незважаючи на це, сумарне скорочення м’яза в цілому в нормальних умовах має одночасний характер. М’язи в організмі в спокої не бувають розслабленими, зберігають напругу, що називається тонус. Причиною тонусу є надходження до м’язів із ЦНС(спинного мозку) нервових імпульсів. Основні функції скелетних м’язів: 1. Скорочення, завдяки чому відбувається переміщення тіла в просторі, забезпечується певна поза тіла; 2. рецепторна функція, бо в м’язах і сухожилках є багато рецепторів: пропріорецептори, хемо -, терморецептори; 3. мімічні м’язи проявляють емоціональний стан людини; 4. м’язи є депо води і солей, тому приймають участь в регуляції водно – сольового обміну в організмі; 5. в м’язах утворюється теплова енергія, тому приймають участь в терморегуляції; 6. в м’язовій тканині відбувається синтез глікогена, АТФ; Сила і робота м 'язів
Скорочуючись, м'язи виконують певну роботу навіть у тому випадку, коли людина знаходиться у спокої. В цих умовах м'язи діють на суглоби нашого тіла, підтримуючи певне положення. Види роботи: динамічна та статистична робота Для обчислення динамічної роботи величину піднятого м'язом вантажу множать на висоту підйому. При виконанні цієї роботи необхідно враховувати правило середніх навантажень: максимальна робота буває лише при середніх навантаженнях, а при не значних навантаженнях і при надмірних величинах робота зменшується. Робота, при якій м'язи скорочуються, не змінюючи своєї довжини, а лише збільшуючи напругу (ізометричну), називається статичною. Наприклад, утримання вантажу в певному положенні. Ця робота визначається множенням піднятої ваги на час її утримання. Робота, яка виконується м'язом, залежить від його сили
Сила м’язів Розрізняють максимальну та абсолютну силу. Максимальну силу визначають максимальним вантажем, який здатен підняти м'яз. Ця сила знаходиться в прямо пропорційній залежності від поперечного перерізу м'яза. Але фізіологічний поперечний переріз (сума поперечних перерізів усіх м'язових волокон) співпадає з геометричним лише в м'язах з поздовжнім ходом волокон, а в м'язів і с косим розтушуванням волокон сума поперечних перерізів кожного волокна може значно перевищувати геометричний поперечний переріз. Тому м'язи с косо розташованими волокнами сильніші за м'язи з повздовжніми волокнами. Для порівняння сили м'язів вираховують абсолютну силу —це відношення максимальної сили до площі фізіологічного перерізу. Наприклад, абсолютна сила литкового м'яза - 5, 9 кг\см жувального 10 кг\см, триголового м'яза плеча - 16, 8кг\см2. Втома м’язів.Поняття про активний відпочинок Тимчасове зниження працездатності в результаті роботи називають втомою. Для пояснення механізмів розвитку втоми було висунуто декілька теорій. За теорією «виснаження», втома розвивається внаслідок втрати м'язом енергетичних запасів - глікогену, АТФ, креатинофосфату, які потрібні для його скорочення. Теорія «засмічення» базується на тому, що в процесі роботи у м’язі накопичуються продукти метаболізму (фосфорна, молочна кислота), які знижують його працездатність. Теорія «удушення» пояснюється розвиток втоми внаслідок гіпоксії (зменшення кисню), яка виникає в процесі роботи. В цілому організмі втома залежить не лише від процесів, що проходять у м’язах, але й від процесів в нервовій системі, яка регулює м'язову активність. Дослідженнями І. М. Сєченова, І.П. Павлова, М. Є. Введенського і О. О. Ухтомського було доведено, що в тривалому збереженні працездатності і розвитку втоми головну роль відіграє центральна нервова система. М.Є. Введенський спостерігав у досліді втому м'яза при подразненні одного з відцентрових нервів. Після цього наносили подразнення іншому відцентровому нерву. На це подразнення м'яз відповідав скороченням з попередньою силою. Очевидно, що втома настає спочатку не в м'язі, а в центральній нервовій системі (периферичні нерви практично невтомні). Вплив вегетативної нервової системи (симпатичних нервів) на роботу м'язів було показано на класичному досліді Орбелі і Гінецинським. Експеримент проводився на нервово-м'язовому препараті жаби. Електричним струмом подразнювали спочатку руховий нерв литкового м'яза; при цьому м'яз скорочувався до того часу, поки не наставала втома. Після цього подразнювали симпатичний нерв, що іннервував цей м’яз, і спостерігали відновлення його працездатності. Таким чином, зробили висновок, що симпатичний відділ вегетативної нервової системи відіграє трофічну роль і збільшує інтенсивність обмінних процесів у м'язі, що призводить до підвищення його працездатності. Без сумніву, кора великих півкуль має вплив на працездатність і розвиток втоми. В експерименті, коли досліджуваному, який виконував значну роботу, навіювалось, що він виконує легку роботу, затрата енергії зменшувалась, а інтенсивність роботи не знижувалась, і втома наставала пізніше. Поширена думка, що найкращим засобом відновлення працездатності є цілковитий спокій. Але дослідження І.М. Сєченова довели, що працездатність відновлюється швидше при активному відпочинку, тобто зміні одного виду діяльності іншим.
Енергетика м 'язового скорочення. Фази теплоутворення. У процесі м'язового скорочення відбувається теплоутворення. Фази теплоутворення: І фаза - початкове теплоутворення включає: а) тепло активації (під час латентного періоду); б) тепло вкорочення (під час скорочення м'яза); с) тепло розслаблення (під час розслаблення м'яза). Це тепло утворюється переважно за рахунок анаеробних хімічних процесів у м'язі. ІІ фаза - запізніле теплоутворення, яке в 1000 разів більше, ніж початкове і триває декілька хвилин, після того як м'язове скорочення закінчилось. 90% тепла протягом цієї фази утворюється за рахунок процесів окислення. Таким чином, утворення тепла у м'язі пов'язано з хімічними процесами, що перебігають без, або за участю О2.
Анаеробні пронеси: -відбувається розпад АТФ —► АДФ + Н3РО4 + Q. Q використовується на забезпечення м'язового скорочення. Запаси АТФ невеликі, тому відбувається постійний ресинтез за рахунок енергії, що утворюється в результаті розпаду; - креатфосфат - креатини + Н3Р04 + 0; - розщеплення глікогену (анаеробний гліколіз) -продукти гліколізу молочна піровиноградна кислота накопичуються у м'язі. Аеробні процеси: полягають в окисленні метаболітів білків, вуглеводів, жирів на мітохондріях. (Q, яке утворюється, використовується на ресинтез АТФ, глікогену).
Властивості гладких м 'язів. Автоматія
Структурною одиницею їх є м'язова клітина (міоцит). Міозит веретеноподібної форми, розмір 20-400мкм. Між собою клітини з'єднані колагеном. Скорочувальний апарат складається з міофібрил, що містить переважно актин. З фізіологічної точки зору міоцити поділяють на: а) поодинокі: локалізовані у структурах ока (радужка, війкові м'язи, артеріоли сім'яних протоків, навколо волосяних цибулин); б) об'єднані у функціональний синцитій: мембрани клітин місцями прилягають одна до одної, утворюючи вставні диски. ПД через нексуси поширюється від однієї м'язової клітини на іншу. Завдяки цьому окремі клітини утворюють функціональний синцитій. До гладких м'язів належать м'язи внутрішніх органів, кровоносних судин. 1. Фізіологічні властивості гладеньких м’язів відрізняються від властивостей скелетних м'язів тим, що вони менш збудливі. 2. Збудження по гладких м'язах проводиться повільніше, ніж по скелетних. їх скорочення розвивається також повільніше і зберігається протягом тривалого часу. 3. Для того, щоб викликати збудження гладкого м'яза, необхідно діяти подразником більшої сили. 4. Характерною особливістю гладких м'язів є їх здатність до спонтанної автоматичної діяльності, тобто здатність скорочуватись під впливом імпульсів, які виникають в них самих. Спонтанні скорочення спостерігаються при дослідженні гладких м'язів шлунка, кишок, нирок, жовчного міхура. Якщо денервовані стрічки цих органів помістити в теплий розчин Рінгера, насичений киснем, вони здатні тривалий час автоматично скорочуватись. 5. Гладкі м'язи за св
|



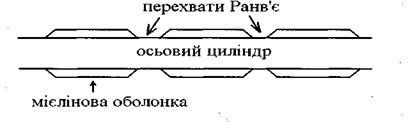 Мієлінове волокно (схема)
Мієлінове волокно (схема)










 Саркомер – структурно-функціональний елемент скорочувального апарату скелетних м’язів. Вони утворені пучками міофібрил, які відокремлені один від одного перпендикулярними смугами – Z-лініями. До Z-ліній прикріпляються одним своїм кінцем тонкі актинові нитки. Інші кінці актинових ниток спрямовані до центру саркомера і входять у проміжки між товстими міозиновими нитками. Частина саркомера, яка примикає до Z лінії та утворена тільки актиновими протофібрилами, має назву І-дисків (ізотропних); слідом за ними розташовані А-диски (анізотропні) – частина саркомера, де має місце взаємне перекриття актинових та міозинових протофібрил. При вкороченні м’яза, коротшає довжина протофібрил і відбувається вкорочення довжини кожного саркомера. Але при цьому довжина анізотропних дисків не зменшується, а зменшується довжина ізотропних дисків. Це є наслідком ковзання актинових протофібрил відносно міозинових в напрямку до центру саркомера.
Саркомер – структурно-функціональний елемент скорочувального апарату скелетних м’язів. Вони утворені пучками міофібрил, які відокремлені один від одного перпендикулярними смугами – Z-лініями. До Z-ліній прикріпляються одним своїм кінцем тонкі актинові нитки. Інші кінці актинових ниток спрямовані до центру саркомера і входять у проміжки між товстими міозиновими нитками. Частина саркомера, яка примикає до Z лінії та утворена тільки актиновими протофібрилами, має назву І-дисків (ізотропних); слідом за ними розташовані А-диски (анізотропні) – частина саркомера, де має місце взаємне перекриття актинових та міозинових протофібрил. При вкороченні м’яза, коротшає довжина протофібрил і відбувається вкорочення довжини кожного саркомера. Але при цьому довжина анізотропних дисків не зменшується, а зменшується довжина ізотропних дисків. Це є наслідком ковзання актинових протофібрил відносно міозинових в напрямку до центру саркомера.
 Причиною руху актинових ниток відносно міозинових (їх ковзання) є " нахил" голівки після її приєднання до активного центру актину. Велика кількість голівок, що входять до складу актоміозинових містків, одночасно роблять " нахил" і нитка актину переміщується до центру саркомера вздовж нитки міозину. За рахунок одного такого " нахилу" саркомер (м'яз) може вкоротитися на 1% своєї вихідної довжини. Ступінь вкорочення м'яза може досягати 50% і такі " нахили" голівок під час одного м'язового скорочення мають повторитися 50 разів.
Причиною руху актинових ниток відносно міозинових (їх ковзання) є " нахил" голівки після її приєднання до активного центру актину. Велика кількість голівок, що входять до складу актоміозинових містків, одночасно роблять " нахил" і нитка актину переміщується до центру саркомера вздовж нитки міозину. За рахунок одного такого " нахилу" саркомер (м'яз) може вкоротитися на 1% своєї вихідної довжини. Ступінь вкорочення м'яза може досягати 50% і такі " нахили" голівок під час одного м'язового скорочення мають повторитися 50 разів. Механізм передачі збудження через нервово-м'язовий синас
Механізм передачі збудження через нервово-м'язовий синас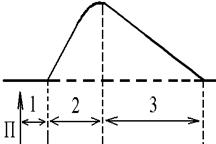 - латентий період (1) - інтервал часу від дії подразника (П) до початку скорочення;
- латентий період (1) - інтервал часу від дії подразника (П) до початку скорочення;



