Р О З В И Т О К З А Р О Д К А.
Д Р О Б Л Е Н Н Я. Зигота зразу ж ділиться на два бластомера, які залишаються з’єднаними один з одним. Далі вони продовжують швидко ділитися, і хвилі клітинних поділів слідують одна за одною майже без перерви. Клітини не встигають збільшитися в розмірах, і велика зигота перетворюється на щільно поєднану масу клітин у вигляді кульки майже такого ж розміру, як була зигота. Тому її поділ називається дробленням. Морфологія дроблення відрізняється у різних хребетних. Один з головних чинників - кількість жовтка, що міститься в клітині. Оліголецитальні клітини діляться повністю (голобластично) майже на однакові бластомери. Таке дроблення, характерне, наприклад, для примітивних хребетних (ланцетника), називається повним і рівномірним. Для мезолецитальних яєць хрящових риб, земноводних властиве повне, але нерівномірне дроблення. При такому виді дроблення утворюються різні за величиною бластомери, причому перших два дроблення дають майже однакові клітини, а потім розташовані на анімальному полюсі діляться швидше, стають дрібнішими, ніж на вегетативному. За розміщенням жовтка яйцеклітини цих тварин телолецитальні, і жовток на вегетативному полюсі яйця гальмує процес дроблення (Рис. 4). Велика маса жовтка в яйцеклітинах плазунів і птахів робить серйозний вплив на дроблення. Цей процес обмежується невеликою ділянкою цитоплазми на анімальному полюсі у вигляді диска, а решта яйцеклітини, багата на жовток, не ділиться. Таке дроблення називається неповним (меробластичним, дискоїдальним). Є і інші види неповного дроблення, наприклад, у членистоногих поверхневе.
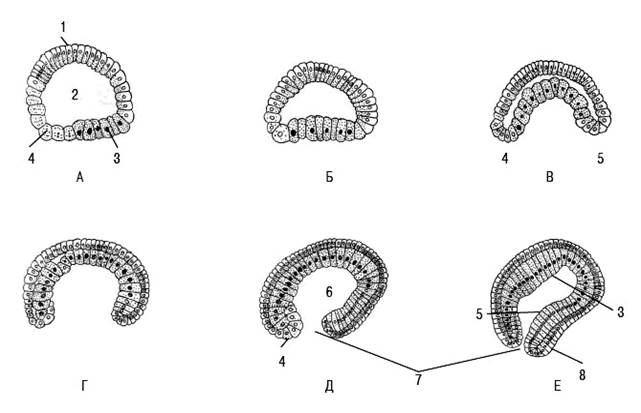
Рис. 4. Схема повного рівномірного дроблення ланцетника (А, І-VII) з утворенням целобластули (Б): Б: 1-бластодерма; 2-бластоцель; 3-дно; 4-крайова зона; 5-дах бластули.
Б Л А С Т У Л А, або одношаровий зародок. Внаслідок дроблення виникає перший багатоклітинний зародок у вигляді кульки - бластула. У найпростішому вигляді у ланцетника всередині кульки є порожнина - бластоцель (первинна порожнина тіла), а стінка його називається бластодермою. Її клітини майже однакового розміру, тільки трохи крупніші на колишньому вегетативному полюсі (дні бластули) в порівнянні з анімальним (дахом бластули). Така типова, правильна бластула називається целобластулою. При дробленні мезолецитальних яєць жаби бластомери вегетативного полюса, навантажені жовтком, крупніші в порівнянні з клітинами анімального полюсу. Тому бластоцель зміщується до даху. Окрім меридіональних і екваторіальних або широтних борозен дроблення у земноводних з'являються і тангенціальні борозни (що йдуть паралельно до поверхні), тому стінка бластули складається з декількох рядів клітин. Такий зародок називається амфібластулою (Рис. 5). При дискоїдальному дробленні зародок має вид диска, розташованого на неподіленому жовтку. Це дискобластула плазунів, птахів. Є і інші види бластул у членистоногих -
перібластула, стерробластула. Рис.5. Схема повного нерівномірного дроблення амфібій (А,I-VI) з утворенням амфібластули(Б): Б-1-мікромери; 2-макромери; 3-бластодерма;4-бластоцель.
Г А С Т Р У Л А, або двошаровий зародок. В процесі гаструляції (утворення гаструли) з бластодерми різними способами утворюються два шари клітин (зародкові листки): зовнішній (ектодерма) і внутрішній (ентодерма). Потім з'являється і третій – середній зародковий листок (мезодерма). Таким чином, відбувається переміщення, диференціювання зовні однакових клітин бластодерми в абсолютно різні за умов існування і функцій зародкові листки, з яких в майбутньому утворюються різні тканини, органи (Рис. 6).
Рис. 6. Типи гаструляції(схема): 1-інвагінація; 2-епіболія; 3-іміграція; 4-деламінація.
Є чотири типи гаструляції: і н в а г і н а ц і я (впинання), е п і бо л і я (обростання), і м і г р а ц і я (вселення), д е п я м і н а ц і я (розшарування). Залежно від кількості жовтка, умов розвитку зародка, виду бластул спостерігається комбінування типів гаструляціі або домінування одного з них. Так, у ланцетника впинання починається від крайової зони (на межі між дахом і дном). Дно бластули прогинається всередину бластоцеля і досягає даху. Після закінчення інвагінації зародок стає двошаровим, з ектодермою і ентодермою. Порожнина називається гастроцель (порожнина первинної кишки), а отвір, що сполучає гастроцель із зовнішнім середовищем, називається бластопор (первинний рот). Він має чотири губи: дорсальну, вентральну і дві бокові (Рис. 7). У амфібій у зв'язку із збільшенням кількості жовтка, невеликим розміром бластоцеля гаструляція з допомогою тільки інвагінації недостатня. Тому активні клітини даху бластули, розмножуючись, оточують крупні клітини дна і утворюють ектодерму, а клітини, навантажені жовтком, дадуть ентодерму. Таким чином, гаструляція у костистих риб, земноводних відбувається при поєднанні двох способів - інвагінації і епіболії. Іміграція включає переміщення клітин бластодерми всередину бластоцеля. Вони осідають на внутрішній поверхні бластодерми і утворюють ентодерму.
Деламінація є розщепленням бластодерми у вигляді диска або щитка на два листки. Рис. 7. Схема гаструляції ланцетника (А-Е): 1-матеріал ектодерми; 2-бластоцель; 3-матеріал ентодерми; 4-матеріал мезодерми; 5-матеріал хорди; 6-гастроцель; 7-бластопор; 8-дорсальна губа бластопору Первинний рот (бластопор) має неоднакову долю у різних тварин в процесі еволюції. Частіше він заростає. Потім у одних тварин на його місці проривається ротовий отвір, а у інших - анальний (задній прохід). Залежно від того, чи відповідає первинний рот зародка остаточному роту або анальному отвору дорослого організму, всі тварини діляться на первинноротих (черв'яки, молюски, членистоногі) і вторинноротих (деякі безхребетні і хордові). Мезодерма (середній зародковий листок) в процесі ембріонального розвитку утворюється різними способами: телобластичним, ентероцельним, ектодермальним і змішаним. У первинноротих матеріал майбутньої мезодерми концентрується в особливих бластомерах - телобластах, тобто виділяється ще в процесі дроблення. У вторинноротих утворення мезодерми тісно пов'язане з ентодермою: вона має вигляд бічних виростів кишкової стінки (ентероцельно). У амфібій вона виділяється в області бічних губ бластопора і в процесі гаструляції одночасно з впинанням ентодерми вона проростає між екто- і ентодермою. У вищих хребетних після утворення гаструли мезодерма тимчасово концентрується в первинній ектодермі, з якої виселяються клітини і розміщуються між зовнішнім і внутрішнім зародковими листками.
Н Е Й Р У Л А. Утворення осьових органів, диференціювання мезодерми. Наступним етапом в розвитку зародка є утворення осьових органів (нервової трубки, хорди, кишкової трубки) і диференціювання мезодерми (первинне - на соміти і спланхнотом і вторинне - подальше їх розділення). Нервова трубка як зачаток нервової системи закладається з ектодерми. На майбутній дорсальній поверхні зародка частина ектодерми утворює медулярну, або нервову пластинку з більш високих клітин. Середня частина її прогинається, а краї мають вигляд виступів. При подальшому розмноженні клітин ці виступи поступово стуляються, утворюється нервова трубка. У хребетних тварин передня частина нервової трубки розширяється, проходить стадію мозкових пухирів, з яких в подальшому розвивається головний мозок. Ектодерма поступово обростає зачаток нервової трубки і зростається над ним. Для всіх хордових характерне утворення спинної струни - хорди, що лежить під нервовою трубкою і має вигляд суцільного клітинного тяжа. Це осьова опорна структура тіла, яка вказує на розташування майбутнього хребта; крім того, під її дією формується нервова система. Більшість вчених вважає, що хорда - один з компонентів мезодерми. У ланцетника вона спочатку має вид пластинки, яка потім завертається в циліндричний тяж і відділяється від ентодерми. Нижче розташована ентодерма округляється і утворює первинну кишку. Мезодерма, розташована з боків, між зачатками осьових органів (нервової трубки, хорди, кишкової трубки) і шкірною ектодермою, вже у амфібій диференціюється схоже з іншими класами хребетних, тому зручніше розглянути ці процеси тут (Рис. 8).
Рис. 8. Гаструла і формування органів у ланцетника (схема):
ект-ектодерма; ент-ентодерма; пк-первинна кишка; нп-нервова плас-тинка; нт-нервова трубка; х-хорда; мез-мезодерма; ц-целом.
П е р в и н н е диференціювання мезодерми, розташованої дорсально, полягає у тому, що вона поступово сегментується по довжині зародка. Ці спинні сегменти називаються сомітами. Вентрально розташована мезодерма (біля первинної кишки) залишається несегментованою і називається бічною пластинкою, або спланхнотомом. Соміти з'єднуються із спланхнотомом звуженими ділянками - сегментними ніжками (нефрогонадотомами).
В т о р и н н е диференціювання полягає перш за все у виділенні з мезодерми першої тканини зародка - мезенхіми (клітин зірчастої форми). В першу чергу це відбувається у вентро-медіальній частині соміта біля хорди, яка називається склеротом. З нього розвивається осьовий скелет. Дорсо-латеральна частина соміта, що залишилася, утворює шкірно-м'язовий пласт. Зовнішня його частина (дерматом) теж дає мезенхіму - зачаток дерми. Середня частина пласта, яка лежить між дерматомом і склеротомом, сильно розростається і дає міотом - джерело розвитку скелетної мускулатури. Спланхнотом розщеп-люється на два листки: зовнішній, пристінковий (парієтальний) і внутрішній (вісцеральний) біля ентодерми. Між листками виникає вторинна порожнина тіла - целом, у вищих тварин це майбутні грудна і черевна порожнини. Разом з утворенням осьових органів і вслід за ними у зародка закладаються зачатки і інших органів і тканин, тобто відбувається органогенез і гістогенез (Рис. 9).
Рис. 9. Стереограма зародка амфібій (схема):
ект-ектодерма; мез-мезодерма; вл-вісцеральний листок мезодерми; пл-парієтальний листок мезодерми; киш-кишечник; нт-нервова трубка; х-хорда; сегм-сегментна ніжка; дерм-дерматом; скл-склеротом; мнх-мезенхіма; спл-спланхнотом; мі-міотом; с-соміт.
П О Х І Д Н І З А Р О Д К О В И Х Л И С Т К І В. У розвитку органів беруть участь два і більш різнорідних зачатків. Кожна тканина виникає з одного ембріонального зачатка в результаті специфічного диференціювання його клітин. Диференціація (розчленовування, розподіл) організму зародка на все більшу кількість різнорідних частин (органів, тканин, клітин) супроводжується інтеграцією, тобто все більш тісним об'єднанням частин в один організм. Виходячи з цього, перерахувати джерела розвитку всіх органів важко, тому розглядаються основні. Е к т о д е р м а дає нервову трубку, тобто зачатки головного і спинного мозку; шкірну ектодерму, тобто епідерміс шкіри і його похідні. Е н т о д е р м а є джерелом утворення епітеліального шару шлунково-кишкового тракту і залоз, органів дихання і їх залоз. М е з о д е р м а має найбільшу кількість похідних: хорду, соміти, нефрогонадотоми, мезенхіму. Від хорди у дорослих тварин залишаються міжхребцеві диски. З міотомів сомітів розвивається скелетна мускулатура, з мезенхіми та дерматомів – сполучнотканинна частина шкіри. Склеротом - джерело розвитку хрящової і кісткової тканин. З мезенхіми в цілому розвиваються тканини внутрішнього середовища (кров, пухка і щільна сполучні тканини і ін). Сегментні ніжки беруть участь в розвитку сечо-статевої системи. З парієтального і вісцерального листків спланхнотома розвиваються, відповідно, пристінковий і внутрішній листки плєври, очеревини; серозні оболонки внутрішніх органів, кіркова речовина наднирника, м'яз серця, фолікулярний епітелій статевих органів і ін.
|