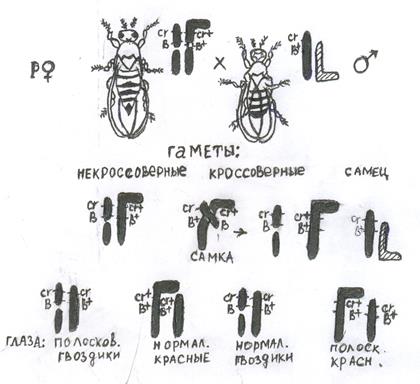
Цитологическое доказательство перекреста хромосом.
После того как было доказано явление кроссинговера генетическими методами, необходимо было получить прямое доказательство обмена участками гомологичных хромосом, сопровождающегося рекомбинацией генов. К. Штерну удалось получить у самки и самца дрозофилы половые хромосомы, отличаемые друг от друга цитологически и генетически, различающиеся по морфологии. У самок отличие х-хромосом получено за счет того, что к одной из них был присоединен фрагмент от у-хромосомы, что дало ч-образную х-хромосому с генами cr B+ дикого типа. Другая х-хромосома была также составной: один из ее фрагментов, несущий центросому, содержал рецессивный ген cr (carnation) – глаза цвета гвоздики и доминантный ген В (carnation – определяет глаза цвета красной гвоздики, а доминантный к дикому типу ген Вач – полосковидные глаза). Безцентромерный фрагмент х-хромосомы не был потерян в силу того, что он прикрепился к IV хромосоме. Так как прибавка фрагмента от у-хромосомы генетически мало активной, вообщем не изменила хромосомного балланса зиготы, гетерозигота по данным генам самка была вполне жизнеспособна. Для анализирующего скрещивания были взяты самцы с нормальными х- и у-хромосомами при этом половая х-хромосома самца несла гены cr и В+ в гемизиготном состоянии. В потомстве были изучены цитологически только самки, среди которых возникли два класса мух с некроссоверными хромосомами crВ и cr+В+ == === crВ+ cr В+ и два других класса мух crВ+ и cr+В == == crВ+ crВ+ самок могли появиться только при условии произошедшего кроссинговера между генами cr и В.
Так как каждый из четырех классов отличался друг от друга не только по признакам глаз, но и по морфологии половых хромосом, то цитологически можно было установить, что кроссинговер, давший два класса мух сопровождался обменом участков х-хромосом. К.Штерн проверил цитологически 374 самки из описанного скрещивания, 369 из них соответствовали ожиданному результату. Подобный же опыт проведен был на кукурузе. В потомстве получили как некроссоверные, так и кроссоверные зерна. У большинства изученных животных и растений хиазмы и перекрест осуществляются у обоих полов. Но существуют отдельные виды животных, у которых кроссинговер осуществляется только у гомогаметного пола, а у гетерогаметного в норме отсутстует. У самцов дрозофилы и самок шелкопряда – полностью отсутствует синапсис, т.е. парное соединение гомологичных хромосом. У гомогаметных особей этих видов (самок дрозофилы и самцы шелкопряда) перекрест хромосом протекает нормально. У многих видов млекопитающих, птиц, рыб и насекомых гетерогаметность пола не сказывается на процессе кроссинговера. Накопленные факты, говорят о том, что в механизме перекреста хромосом большую роль играет центромера. Вблизи центромеры перекрест происходит редко, затем у дрозофилы возрастает, а к концу снова уменьшается. Поэтому на генетических картах дрозофилы вблизи центромеры локализуется генов больше, чем у удаленных от нее районах. У кукурузы такого влияния центромеры на кроссинговер не обнаружено. На частоту перекреста в разных участках хромосомы влияет и распределение гетерохроматиновых и эухроматиновых участков. У гетерохроматинового пола (самцы дрозофилы и самки тутового шелкопряда) перекрест хромосом не обнаруживается. Однако, если эти организмы подвергнуть действию рентгеновских лучей, то в потомстве возникают кроссоверные особи. Частота кроссинговера также зависит от возраста организма. Если изучить перекрест между двумя генами, локализованными по генетической карте на небольшом расстоянии, скажем на растоянии 6 морганид друг от друга, и учитовать процента кроссинговера у самок по десяти дневкам откладки оплодотворенных яиц (за 1-10, 11-20, 21-30 дней), то частота перекреста будет колебаться соответственно: 5,9; 1,8; 3,8%. Первый возраст соответствует максимому, второй – спаду, а третий – подъем процента перекреста. У генетика не вызывает сомнения положение о том, что все механизмы, работающие в клетке организма, наследственно детерменинированы. Кроссинговер тоже контролируется генотипом. Подтверждением этому служит эффективность отбора линий на высокую и низкую частоту перекреста между двумя генами. Отбор линий по данному признаку указывает на роль генотипа в определении кроссинговера. На частоту кроссинговера генотип может влиять разными путями. На молекулярном уровне генотип регулирует точность копирования ДНК. На хромосомном уровне генотип влияет через изменение политении хромосом, степень спирализации, прочность продольных связей в нити ДНК. На частоту перекреста в сильной степени влияют различные хромосомные перестройки, поскольку они нарушают нормальный синапсис хромосом. Имеется и межхромосомное влияние на перекрест. Хромосомные перестройки произошедшие в одной паре хромосом, влияют на частоту перекрестта в негомологичных хромосомах. У кукурузы открыты гены, которые контролируют синапсис, спирализацию и слияние хромосом. Помимо зависимости кроссинговера от генотипа и физиологического состояния, на частоту перекреста оказывают влияние факторы среды: высокая и низкая температуры, ионизирующее излучение, инфракрасные лучи и т.д. Вопросы для самоконтроля: 1.Что такое кроссинговер. 2.Цитологическое доказательство кроссинговера. 3.Генетическое доказательство кроссинговера. Рекомендуемая литература: М.Е. Лобашев и др.«Генетика с основами селекции». М., МГУ, 1978год. 425стр. С.Г. Инге-Вечтомов «Генетика с основами селекции». Москва «Высшая школа». 1989год, 590стр. Р.Г. Заяц. и др. «Общая и медецинская генетика». Ростов- на- Дону. «Феникс». 2002год. 315стр.
|