ЧАСТЬ I. ОБЩАЯ ФИЗИОЛОГИЯ КЛЕТКИ
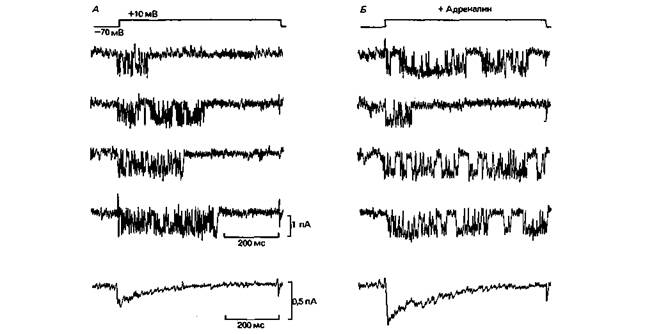
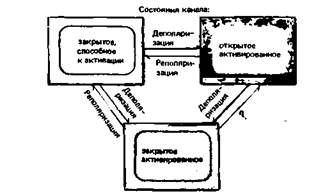
Токи одиночных Ca2 +-каналов в миокарде (рис. 2.14) характеризуются несколько более сложным поведением по сравнению с Na+- и К+-токами, показанными на рис. 2.12. Во время серий деполяризационных скачков потенциала примерно в 70% случаев возникают довольно длительные вспышки импульсов тока, каждый амплитудой около 1 пА, а в 30% случаев канал остается закрытым. Индивидуальные открывания во время вспышек продолжаются в среднем около 1 мс, а закрытые состояния между ними - только 0,2 мс. Суммарный Ca2+-ток во время деполяризации (нижние записи на рис. 2.14) быстро нарастает и инактивируется с постоянной времени примерно 130 мс, причем общий ток определяется длительностью и частотой вспышек. Кинетику канала проще всего описать (в соответствии с рис. 2.13) следующим уравнением: Закрытое состояние 1
Здесь от переходов между «Закрытым состоянием 2» и «Открытым состоянием» зависит длительность и частота индивидуальных открываний, а от переходов между «Закрытым состоянием 1» и «Закры-
ГЛАВА 2. ПЕРЕДАЧА ИНФОРМАЦИИ ПОСРЕДСТВОМ ВОЗБУЖДЕНИЯ 39 тым состоянием 2»-частота и длительность вспышек. Уравнение (2) требует дополнения, чтобы учесть инактивированное состояние, как показано на рис. 2.13 [32]. Записи активности Ca2 +-канала на рис. 2.14 служат также примером модуляции активности канала гормоном или медиатором (см. с. 65). Адреналин,секретируемый корой надпочечников как «эрготропный гормон», поступает к сердцу с кровотоком; один из его эффектов состоит в увеличении частоты сердечных сокращений. Кроме того, он высвобождается (вместе с норадреналином) в качестве медиатора из симпатических нервов сердца, вызывая тот же эффект (с. 462). В эксперименте, результаты которого приведены на рис. 2.14, Б, адреналин в концентрации 10 −6Μ апплицировали на клетку миокарда. После этого деполяризация вызвала примерно в 80% случаев активность одиночных Ca2+-каналов с повышенной частотой вспышек. Кратковременные открывания и закрывания каналов были такими же, как раньше. Суммарная кривая (рис. 2.14, Б. внизу) отчетливо показывает, что адреналин увеличивал вход Са2 +. Такой же эффект можно вызвать перфузией клеток миокарда раствором с циклическим аденозинмонофосфатом (цАМФ) или применением каталитической субъединицы цАМФзависимой протеинкиназы (ПК-Α). Эти наблюдения свидетельствуют, что адреналин действует здесь через второй посредник -цАМФ, вызывая фосфорилирование ферментов каталитической субъединицей протеинкиназы (рис. 1.15, с. 24) [19]. Таким образом, адреналин, по-видимому, увеличивает Ca2+-ток путем инициации фосфорилирования Ca2 +-канала, которое способствует переходу из «Закрытого состояния 1» в «Закрытое состояние 2». Эффект адреналина, представленный на рис. 2.14, может служить прототипом модуляции клеточной активности гормонами или медиаторами. В мембране, несомненно, существуют еще и Cl− -каналы. Они изучены недостаточно подробно, поэтому рассматриваться здесь не будут. Молекулы Na+-канала. Белки различных каналов очень сходны между собой по структуре и функциям; полагают, что все они происходят от Са2+канала. Поскольку наиболее тщательно исследована молекула Na+-канала, мы вновь обратимся к нему. Na+-канал состоит из гликопротеина с молекулярной массой ~ 300 000. Недавно установлена его аминокислотная последовательность. Изолированные молекулы можно включить в искусственные липидные мембраны, где они продолжают функционировать [8]. Число имеющихся в мембране Na+каналов можно определить путем «титрования» тетродотоксином, который связывается с этими каналами, или путем деления величины Na+-тока через мембрану площадью 1 мкм2 на амплитуду тока одного канала. Разные типы мембран содержат от 1 до 50 каналов на 1 мкм2. При плотности 50 каналов· мкм−2 среднее расстояние между ними составляет около 140 нм. Если принять диаметр молекулы канала равным примерно 8 нм, а диаметр просвета канала, когда он открыт,- около 0,5 нм, то оказывается, что каналы находятся друг от друга довольно далеко. В течение 1 мс открытого состояния через один такой канал входит примерно 1 пА тока, перенося заряд, равный 10−15 Кл. Емкость мембраны обычно равна 1 мкФ см−2 или 10−14 Ф мкм−2. Поскольку 1Ф = 1 Кл-В−1, заряд величиной 10−15 Кл мкм−2. который входит в клетку за время одного открывания каналов, достаточен для смещения мембранного потенциала на 100 мВ; иными словами, такой заряд обеспечивает фазу нарастания потенциала действия. Заряд величиной 10−15 Кл переносит 6000 ионов Na +. Повышение внутриклеточной концентрации, обусловленное поступлением 6000 ионов Na+ в примембранную область объемом 1 мкм3, пренебрежимо мало, 10−5 М. Следовательно, токи каналов достаточно велики для обеспечения генерации потенциала действия, но не создают заметных изменений внутриклеточных концентраций ионов (за исключением [Ca2+]i). Таким образом, восстановление трансмембранных ионных градиентов посредством Na/К-насоса (с. 15) не играет роли в случае одиночного потенциала действия. Белок Na+-канала должен быть способен не только быстро включать массивный поток Na+, но и предотвращать одновременный вход других ионов, особенно К+, которые имеют почти те же размеры. Значит, Na+ -каналы должны характеризоваться избирательностью. Что касается анионов, то они удерживаются отрицательными зарядами у входа в канал, как это показано на схеме (рис. 2.15). Из мелких катионов Li + проходит через Na+-канал относительно хорошо, тогда как К+ практически не пропускается. Избирательность можно объяснить только специфическим связыванием иона во время его прохождения через канал, о чем уже говорилось при обсуждении энергетического уровня связывания вдоль канала (рис. 1.5, Б) [21]. Кроме избирательности для Na+, Na+-канал должен обладать способностью быстро изменять свою проницаемость при изменениях мембранного потенциала. Следовательно, молекула Na+-канала должна нести заряды, которые могут смещаться под влиянием сдвигов силы электрического поля через мембрану. Смещения этих зарядов регистрируются в виде «воротных токов» [3, 9, 23] после полной блокады ионных каналов; воротные токи свидетельствуют о смещении по крайней мере 4 зарядов на канал. Эти 4 заряда представлены на рис. 2.15 как «датчик электрического поля», способствующий изменению конформации молекулы, при котором ка- 40 ЧАСТЬ I. ОБЩАЯ ФИЗИОЛОГИЯ КЛЕТКИ нал открывается. Открытое состояние нестабильно и преобразуется спонтанно в закрытое инактивированное состояние. Инактивация осуществляется участками канального белка, находящимися на внутренней стороне мембраны. Вещества, которые действуют внутриклеточно, например иодат или проназа, а также специфические токсины и фармакологические препараты, могут блокировать инактивацию. Еще один способ блокады Na+-канала представляет интерес для медицины. Местные анестетики используются для предотвращения генерирования и распространения возбуждения в нервах, с тем чтобы потенциалы действия от «болевых рецепторов» не поступали в ЦНС. Анестетики обычно вводят около того нерва, который нужно блокировать. Однако их молекулы связываются только с открытыми каналами, в участке между входом в селективную пору и «воротами» (рис. 2.15) [25. 30]. Молекулы местных анестетиков слишком велики, чтобы войти в устье канала с наружной стороны мембраны. Они могут входить в открытый канал только с внутренней стороны мембраны или же, если они жирорастворимы, через липидную мембрану. Вызываемые ими закрывания канала часто продолжаются только несколько миллисекунд, но повторяются с высокой частотой; разбивая ток одиночного канала на много коротких фрагментов, анестетики делают вход Na+ неэффективным.
|
 Закрытое состояние 2
Закрытое состояние 2 Открытое состояние (2)
Открытое состояние (2)