Биологическое окисление в митохондриях
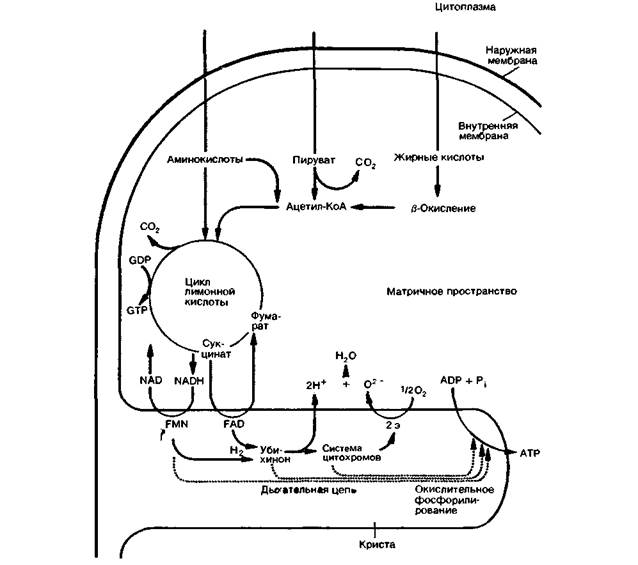
Биологическое окисление происходит в митохондриях. Кроме ферментов цикла лимонной кислоты, дыхательной цепи и окислительного фосфорилирования в этих органеллах были обнаружены также ферменты расщепления жирных кислот и ряда аминокислот [4]. На рис. 23.1 схематично показаны различные пути окислительного метаболизма в митохондриях. Пируват. жирные кислоты и аминокислоты переносятся из цитоплазмы через митохондриальные мембраны в матрикс митохондрий. После ряда биохимических превращений все эти субстраты распадаются до веществ, которые поступают в цикл лимонной кислоты. Пируват, образующийся главным образом в процессе аэробного расщепления глюкозы в цитоплазме, превращается в результате окислительного декарбоксилирования в матриксе митохондрий в ацетилкофермент А (ацетил-КоА), большая часть которого в нормальных условиях распадается в цикле лимонной кислоты. В отличие от расщепления глюкозы, первый этап которого (гликолиз) протекает в цитоплазме, окислительный распад жирных кислот полностью происходит в матриксе митохондрий. Молекулы ГЛАВА 23. ТКАНЕВОЕ ДЫХАНИЕ 627
жирных кислот окисляются в цепи биологических реакций, называемой ß-окислением; образующийся при этом ацетил-КоА поступает в цикл лимонной кислоты либо используется для синтеза жирных кислот. Органические кислоты, образовавшиеся в матриксе митохондрий при метаболизме аминокислот, могут включаться в цикл лимонной кислоты на разных этапах в виде ацетил-КоА, а-кетоглутарата, сукцинил-КоА, фумарата и оксалоацетата. Образовавшиеся в цикле лимонной кислоты молекулы НАДН (NADH, продукт трех реакций дегидрогенизации) и сукцинат диффундируют из матрикса митохондрий к их внутренней мембране, в которой локализованы ферменты дыхательной цепи и окислительного фосфорилирования. Здесь НАДН подвергается окислению комплексом ферментов дыхательной цепи, включающим в качестве кофермента флавинмононуклеотид (FMN). Окисление сукцината осуществляется комплексом ферментов дыхательной цепи, в котором роль кофермента выполняет флавинадениндинуклеотид (ФАД, FAD). Оба флавопротеиновых комплекса передают электроны на убихинон (кофермент Q), который затем окисляется с участием цитохромов. В конечном счете из каждой молекулы водорода образуется 2 протона и 2 электрона. Далее электроны переносятся специфическими железосодержащими компонентами дыхательной цепи (цитохром b, FeS-белок, цитохром с1, цитохром с 2 ) на цитохромоксидазу (комплекс 628 ЧАСТЬ VI. ДЫХАНИЕ цитохромов а и а3), которая передает их на молекулярный кислород. Восстановленный кислород соединяется со свободными ионами водорода с образованием воды. Энергия, высвобождающаяся в процессе переноса электронов по дыхательной цепи, используется для перекачивания протонов из матрикса митохондрий в пространство между их внутренней и наружной мембранами. Перенос протонов приводит к созданию мембранного потенциала на внутренней митохондриальной мембране. За счет тока протонов обратно в матрикс митохондрий индуцируется, согласно теории Митчелла, синтез АТФ (окислительное фосфорилирование). При окислении каждой молекулы НАДН образуются три молекулы АТФ, а при окислении одной молекулы ФАДН2-две молекулы АТФ. В первом случае отношение образования АТФ к потреблению кислорода (коэффициент Р/О) равно 3 (на 1 грамм-атом кислорода образуются 3 моль АТФ), а во втором-2. Последствия недостаточного снабжения тканей кислородом. При ряде патологических состояний нарушается снабжение тканей кислородом. В этих случаях энергетические потребности клеток могут в течение короткого времени удовлетворяться за счет ограниченных запасов энергии в виде АТФ и креатинфосфата, а также за счет анаэробного процесса-гликолиза. Однако этих источников энергии недостаточно, и они могут использоваться лишь в течение небольшого периода времени, что вызвано двумя основными причинами. Во-первых, в анаэробных условиях потребность клеток в глюкозе настолько возрастает, что чаще всего не может полностью удовлетворяться в течение длительного времени. Во-вторых, в процессе гликолиза образуется в больших количествах лактат, который не столь быстро удаляется из ткани для использования в других органах (например, для расщепления в печени, почках или миокарде или для синтеза гликогена). При сильном недостатке кислорода содержание лактата в тканях и крови постоянно возрастает, что в конечном счете приводит к нереспираторному ацидозу. Когда pH внутриклеточной среды падает ниже уровня, оптимального для активности ферментных систем, наступают резкие нарушения в клеточном метаболизме.
|