
Потребность тканей в кислороде
Потребление О2 в состоянии покоя. Количество кислорода, потребляемого тканью, зависит от функционального состояния входящих в ее состав клеток. В табл. 23.1 приведены данные о потреблении кислорода различными органами и их частями, когда организм находится в состоянии покоя при нормальной температуре. Скорость потребления кислорода тем или иным органом ( выражают в мл О2 на 1 г или 100 г массы за 1 мин (при этом учитывается масса органа в естественных условиях). В соответствии с принципом Фика
Когда организм находится в состоянии покоя, кислород относительно интенсивно поглощается миокардом, серым веществом головного мозга (в частности, корой), печенью и корковым веществом почек. В то же время скелетные мышцы, селезенка и белое вещество головного мозга потребляют меньше кислорода (табл. 23.1). Различия в потреблении кислорода разными участками одного и того же органа. Во многих органах можно измерить кровоток через ограниченные участки ткани путем определения клиренса инертных газов (например, 85Кг, 133Хе и Н2). Таким образом, если возможно взять пробу крови из вены, по которой осуществляется отток от данного участка, то этот метод позволяет определить в нем потребление кислорода. Кроме того, несколько лет назад был разработан метод позитронной эмиссионной томографии (ПЭТ), позволяющий непосредственно измерять кровоток и потребление О2 в отдельных частях органов. Этот метод успешно применяется для исследования головного мозга человека [21, 36]. До внедрения метода ПЭТ, как видно из табл. 23.1, измерить региональное потребление О2 можно было лишь в немногих органах. При изучении потребления кислорода тканями мозга различных млекопитающих было показано, что кора больших полушарий потребляет от 8 10−2 до 0,1 мл О2 г−1мин−1. Исходя из потребления О2 целым мозгом и корой, можно вычислить среднее потребление О2 белым веществом мозга. Эта величина составляет примерно 1 10 −2 мл г−1 мин−1. Прямое измерение поглощения О2 участками головного мозга у здоровых испытуемых методом позитронной эмиссионной томографии дало следующие величины: для серого вещества (в различных участках) - примерно от 4 до 6-10−2 мл г−1 -мин−1, для белого вещества-2-10 − 2 млг−1 мин−1 [21, 36]. Можно предполагать, что потребление кислорода варьирует не только в зависимости от участка, но также в разных клетках одного участка. В самом деле, при измерении (с помощью платиновых микроэлектродов) регионального потребления О2 поверхностными клеточными слоями коры головного мозга было показано, что в условиях слабого наркоза это потребление в пределах небольших участков варьирует примерно от 4-10−2 до 0,12 мл-г−1-мин−1. Результаты радиоавтогра- ГЛАВА 23. ТКАНЕВОЕ ДЫХАНИЕ 629
Потребление О2 в условиях повышенной активности органа. В том случае, если активность какоголибо органа по тем или иным причинам повышается, в нем возрастает и скорость энергетического обмена, а следовательно, и потребность клеток в кислороде. При физической нагрузке потребление О2 тканями миокарда может увеличиваться в 3-4 раза, а работающими скелетными мышцами -более чем в 20-50 раз по сравнению с уровнем покоя. Потребление О2 тканями почек возрастает при увеличении скорости реабсорбции Na+. В большинстве органов скорость поглощения О2 не зависит от скорости кровотока в них (при условии, что напряжение О2 в тканях достаточно велико). Почки составляют исключение. Существует критическая скорость перфузии, превышение которой вызывает образование ультрафильтрата; при этом уровне фильтрации повышенный кровоток сопровождается повышенным потреблением О2 тканью почек. Такая особенность обусловлена тем, что интенсивность клубочковой фильтрации (а следовательно, и реабсорбции Na+) пропорциональна скорости кровотока. Зависимость потребления О2 от температуры. Потребление О2 тканями крайне чувствительно к изменениям температуры. При снижении температуры тела энергетический обмен замедляется, а потребность большей части органов в кислороде уменьшается. При нормальной терморегуляции активность органон, участвующих в поддержании теплового баланса, увеличивается, а потребление ими кислорода возрастает. К таким органам относятся, в частности, скелетные мышцы; их терморегуляторная функция осуществляется за счет повышения мышечного тонуса и дрожи (с. 667). Увеличение температуры тела 63β ЧАСТЬ VI. ДЫХАНИЕ сопровождается повышением потребности большинства органов в кислороде. Согласно правилу Вант-Гоффа, при изменении температуры на 10 oС в пределах от 20 до 40oС потребление тканями кислорода изменяется в том же направлении в 2 3 раза (Q10 = 2-3). При некоторых хирургических операциях бывает необходимо временно остановить кровообращение (а следовательно, снабжение органов О2 и питательными веществами). При этом, чтобы уменьшить потребности органов в кислороде, часто используют гипотермию (понижение температуры тела): больному дают такой глубокий наркоз, при котором терморегуляторные механизмы подавляются.
|
 ) обычно
) обычно определяют, исходя из кровотока (
определяют, исходя из кровотока ( ) через тот или иной орган и разницы в концентрациях О2 в поступающей к органу артериальной крови и оттекающей от него венозной крови (
) через тот или иной орган и разницы в концентрациях О2 в поступающей к органу артериальной крови и оттекающей от него венозной крови ( ):
): (1)
(1) ), артериовенозной разницы по О2 (
), артериовенозной разницы по О2 ( ) и потребления 02 (
) и потребления 02 ( ) в различных органах человека при 37 °С
) в различных органах человека при 37 °С








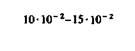
















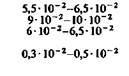





 фических исследований регионального кровотока (с использованием иод-14С-антипирина) и регионального потребления глюкозы (с использованием 14С-2дезоксиглюкозы) в коре головного мозга позволяют считать, что эти параметры также существенно различаются в соседних участках [37, 38]. У людей старше 30 лет региональный кровоток и потребление О2 в сером веществе головного мозга с возрастом постепенно снижаются [19, 21, 36]. Примерно такие же различия в потреблении кислорода были обнаружены между отдельными частями почек. В корковом веществе почек среднее потребление О2 в несколько раз больше, чем во внутренних участках и сосочках мозгового вещества. Поскольку потребности почек в кислороде зависят главным образом от интенсивности активной реабсорбции Na+ из просвета канальцев в ткани, полагают, что столь выраженные различия в региональном потреблении О2 обусловлены в основном разницей между величинами этой реабсорбции в корковом и мозговом веществе [11].
фических исследований регионального кровотока (с использованием иод-14С-антипирина) и регионального потребления глюкозы (с использованием 14С-2дезоксиглюкозы) в коре головного мозга позволяют считать, что эти параметры также существенно различаются в соседних участках [37, 38]. У людей старше 30 лет региональный кровоток и потребление О2 в сером веществе головного мозга с возрастом постепенно снижаются [19, 21, 36]. Примерно такие же различия в потреблении кислорода были обнаружены между отдельными частями почек. В корковом веществе почек среднее потребление О2 в несколько раз больше, чем во внутренних участках и сосочках мозгового вещества. Поскольку потребности почек в кислороде зависят главным образом от интенсивности активной реабсорбции Na+ из просвета канальцев в ткани, полагают, что столь выраженные различия в региональном потреблении О2 обусловлены в основном разницей между величинами этой реабсорбции в корковом и мозговом веществе [11].


