
ОБЩЕСТВА 2 страница
16 Глава первая
Таким образом, информация об изменениях эндокранов в эволюции гоминид и в период перехода от неандертальского человека к современному — это одновременно информация о постепенности этого перехода и продолжении в нем тенденций предшествующего эволюционного развития всего семейства гоминидных предков современного человека. В этой связи особый интерес приобретает проблема так называемого второго скачка в процессе антропогенеза. В советской антропологической и философской литературе наибольшей популярностью пользуется гипотеза двух скачков, двух переходов количественных изменений в качественные: одного — при образовании самого семейства гоминид и второго — при переходе от неандертальца к человеку современного вида30. Аргументировалась и другая точка зрения — о наличии трех скачков (промежуточный скачок между питекантропами и неандертальцами), но она стоит особняком31. Строго говоря, образование любого таксона представляет собою переход количества в качество в ходе эволюционного процесса и, следовательно, возникновение любого вида — эволюционный скачок. Скачок этот тем значительнее, чем более крупный таксон образуется. Выше мы имели возможность убедиться в том, что переход от человека неандертальского вида к современному человеку носил постепенный характер с морфологической точки зрения и с точки зрения тех эволюционных закономерностей, которые управляли этим переходом. Поэтому, никак не отрицая исключительного масштаба культурных преобразований, связанных с возникновением верхнепа-леолитпческого человечества (впрочем, как мы теперь хорошо знаем, истоки многих из них прослеживаются в зачаточных формах в мустьерской культуре), не следует, оставаясь в рамках только антропологической классификации, переоценивать масштабов скачка, падающего на переход от неандертальского к современному виду. Он никак не больше, в свете всего сказанного, а значительно меньше, чем скачок при переходе от питекантропов к неандертальцам. Поэтому же, если говорить о втором скачке в процессе антропогенеза, о втором перерыве постепенности и переходе количественных изменений в качественные, вполне можно помещать его не па заключительном этапе эволюции гоминид, при переходе от Homo neander-thalensis к Homo sapiens, а при образовании рода Homo, при переходе от рода Pithecanthropus к роду Homo32. При этом будут соблюдены все морфологические критерии и последовательность масштаба переходов от одного таксона к другому в рамках антропологической классификации.
______ ЗАВЕРШЕНИЕ ПРОЦЕССА АНТРОПОГЕНЕЗА __________ 17 Возвращаясь к факторам формирования человека современного вида, можно считать очевидным в связи со всем сказанным, что процесс появления Homo sapiens, как и вся предшествующая эволюция гоминид, — процесс полифакторный, и поэтому перечисленные и частично рассмотренные выше гипотезы монофакторной эволюции оставляют необъясненными многие аспекты этого процесса. На протяжении его действовали как общие тепдепцпп эволюции гоминид, которым не вполне правомерно приписывалось ограниченное действие только в пределах этапа перехода от неандертальца к современному человеку, так и специфические тенденции, характерные только для этого этапа. Общие тенденции, как мы помним, — интенсификация социальных аспектов коллективного поведения и общий дальнейший подъем ассоциативного мышления. Высказывалось мнение, что особи с развитыми социальными инстинктами должны были подвергаться преследованиям в обезьяньем стаде и, не выдержав конкуренции с более физически сильными и агрессивными особями, даже погибать33. Вытеснялись они будто бы и из процесса размножения, хотя подобное представление является рецидивом теперь оставленных под давлением фактических данных взглядов о безудержном разгуле «зоологического индивидуализма» в сообществах обезьян и древнейших предков человека. Но основное возражение против подобного представления состоит даже не в этом, было справедливо отмечено, что проблема должна быть перенесена с внутригруппового на межгрупповой уровень: если даже над более альтруистично настроенными индивидумами доминировали агрессивные особи внутри группы (хотя положительная связь между агрессивным поведением п физической силой, жизненной активностью и трудовой ловкостью никогда не была доказана, а лишь постулировалась часто без всяких строгих доказательств), то группы со случайно*! концентрацией особей повышенной социальности приобретали огромное преимущество над первобытными стадами с наличием впутрпстадных конфликтов 34. Специфичные для рассматриваемого этапа частные тенденции эволюции должны объяснить изменение некоторых структурных элементов черепа современного человека по сравнению с неандертальским видом и аналогичное уменьшение массивности скелета. Развитие мозга легко объясняет перестройку лобного отдела черепа, но более прямое положение лобной кости у современного человека не приводит автоматически к уменьшению надорбитного рельефа, примером чему являются черепа неандертальцев группы Схул, особенно череп Схул V: при практически современном наклоне лба этот череп имеет вполне четко выраженный надглазничный валик. Не объясняет прогрессивная перестройка мозга и редукции затылочного рельефа черепа. По-видимому, мы сталкиваемся в данном случае с остаточной, затухающей эволюционной тенденцией, которая погасла после формирования человека современного вида, но начало которой уходит еще в эполюцию приматов.
18 Глава первая
Массивность скелета у подавляющего большинства неандертальских форм сама по себе заслуживает внимания как морфологическое свойство, до какой-то степени выделяющее неандертальский вид среди других видов ископаемых гоминид. Представляется весьма вероятным, что такое общее свойство, охватывающее многие структурные особенности скелета и имеющее большое жизненное значение, могло образоваться в результате специфических условий жизни коллективов неандертальских людей. Была сделана попытка увидеть эту специфику в известном противоречии между расширяющимися возможностями и усовершенствовавшимися способами охоты, с одной стороны, и четко выраженной оседлостью поселений, с другой36. Расширение и дифференциация охотничьих навыков не могли не иметь своим результатом расширение охотничьих территорий и увеличившуюся эффективность охотничьей добычи. Это в свою очередь вело к необходимости перетаскивать добычу на далекие расстояния. Физически сильные невысокие индивидуумы плотного телосложения, с массивным костяком и мощной мускулатурой, безусловно подвергались в этих условиях положительной селекции, что привело в конце концов к наследственному закреплению подобного соматотипа. Учитывая выявленную обширными физиологическими исследованиями значительную роль костного мозга в кроветворении при больших физических нагрузках 37 и продемонстрированную у современного человека отрицательную корреляцию между поперечными размерами и толщиной стенок диафизов длинных костей конечностей, мож-
____________ ЗАВЕРШЕНИЕ ПРОЦЕССА АНТРОПОГЕНЕЗА______________ 19 но предполагать, что массивный костяк у неандертальского человека был еще дополнительно необходим для обеспечения функции гемо - поэза — процесса обновления красных элементов крови. Естественно, и роль отбора па массивность скелета, и его кроветворная функция должны были в связи с изменением условий жизни постепенно падать при переходе к верхнепалеолитическому человеку и далее на протяжении ранних этапов эволюции современного человека. Краткий итог всего изложенного в этом разделе сводится к тому, что, вопреки монофакторным гипотезам происхождения человека современного вида, процесс становления Homo sapiens — процесс полифакторный, в котором слились различные тенденции эволюции, имевшие самостоятельное значение. Эти тенденции, повторяем, — дальнейшая эволюция тех структур мозга и связанных с ними структур черепа, которые обеспечивают повышение уровня ассоциативного мышления и развитие социального поведения, редукция морфологических образований на черепе, потерявших функциональное значение в связи с грацилизацией нижней челюсти и четкой фиксацией выпрямленного положения головы, грацилизация скелета в связи с постепенной потерей им опорных функций при переноске больших тяжестей и активной функции гемопоэза. Только такой подход, учитывающий многие формообразующие факторы, дает возможность сколько-нибудь полно охватить весь процесс становления Homo sapiens в целом и разумным образом объяснить происхождение всей суммы перечисленных в предыдущем разделе морфологических отличий его от человека неандертальского вида. 3. Время формирования Homo sapiens До обоснования А. Хрдличкой неандертальской формы в эволюции современного человека представление об очень раннем отделении эволюционной ветви, ведущей к современному человеку, было всеобщим. В палеоантропологичоской литературе периодически дискутировались находки якобы большого хронологического возраста и не имеющие примитивных признаков. Более тщательное рассмотрение условий их залегания обнаруживало, как правило, неясность их геологической датировки и лишало их серьезного значения для обсуждаемой нами темы38. Но даже и после почти всеобщего принятия концепции А. Хрдлички предположение о возможной глубокой древности человека современного вида не исчезло из арсенала научных теорий, при-мером чему является гипотеза так называемого прссапиенса39. Аргументами в пользу этой гипотезы были две находки — Сванскомб в Англии и Фонтешевад во Франции, которые при заведомо доказанном нижнепалеолитическом возрасте имели недостаточно ясную морфологию, которая позволяла трактовать имеющиеся фрагмепты как принадлежащие человеку современного вида. Все же после критической работы Я. Я. Рогинского 40, Э. Брайтингера41 и Э. Тринкауса42 вопрос о принадлежности черепов из Сванскомба и Фонтешевада к неан-
20 Глава первая
Оживление интереса к этому вопросу связано с находками черепов сравнительно раннего хронологического возраста в долине Омо в Эфиопии, получивших в литературе обозначения Омо I и Омо II43. Первые оценки возраста в 60 000 лет вызвали в дальнейшем сомнения 14, что имеет принципиальное значение, так как морфологическая характеристика обоих черепов недостаточно ясна. Очень похоже, что они различаются на видовом уровне и череп Омо I должен быть отнесен к современному виду, а череп Омо II имеет примитивные особенности. Так или иначе эти находки недостаточно выразительны, чтобы отказаться от уже утвердившегося мнения об эволюционной последовательности двух видов. Можно лишь предположить, что они сосуществовали на каком-то очень коротком отрезке четвертичной истории, когда современный вид формировался в недрах неандертальского. Подтверждение этого — обнаружение скелета ребенка современного вида в позднемустьерских слоях стоянки Ста-роселье в Крыму. При первом описании находки были отмечены на ней отдельные примитивные особенности45, которые не получили подтверждения в дальнейших исследованиях46. С. Н. Замятины высказал сомнения в синхронности скелета с позднемустьерским слоем, из которого он происходит 47, но, по-видимому, сомнения эти неосновательны 4S. Дополнительную сложность в истолкование этой находки вносит своеобразие ее морфологии, носящее, возможно, частично патологический характер: восстановление ее «взрослых» размеров, оправдавшее себя во многих других случаях, дало крайне странные соотношения, не укладывающиеся в рамки современного вида49. Но подобное своеобразие не отменяет ее видовой диагностики и принадлежности к современному виду. Если опираться только на местонахождения с бесспорной стратиграфией и достаточно точными определениями абсолютного возраста, можно констатировать, что остатки человека современного вида не уходят глубже, чем на 40 000 лет. 4. Морфологическая типология и локальная дифференциация верхнепалеолитического человечества Уже первые находки черепов верхпепалеолптического времени, сделанные еще в прошлом веке, описывались не только с чисто морфологической стороны и выраженности на них эволюционно примитивных признаков, но и под углом зрения близости к тем или иным морфологическим вариантам внутри современного человечества. Че-
_____________ ЗАВЕРШЕНИЕ ПРОЦЕССА АНТРОПОГЕНЕЗА_______________ реп из Шанселяда (Франция) долгие годы сближался с эскимосскими черепами5Q, и только реконструкция носовой области позволила показать его четкие отличия от монголоидных серий и, наоборот, сходство с европеоидами51. Аналогичным образом черепа из грота Детей в Гримальди (Италия) — один женский и второй, принадлежавший неполовозрелому субъекту, возможно, юноше — рассматривались как негроидные52, пока повторная обработка материала не показала дефектность первоначальной реконструкции и позволила обнаружить значительное сходство этих черепов с другими европейскими верхнепалеолитическими находками 53. В дальнейшем, по мере увеличения числа известных находок верхпепалеолитического времени и накопления опыта их интерпретации, было предложено несколько схем их морфологической типологии и генетических взаимоотношений, опиравшихся в основном на европейский материал, более многочисленный и лучше изученный54. Теоретически говоря, малоперспективны обе крайние тенденции в оценке морфологической типологии верхнепалеолитического человечества — видеть в нем единую комбинацию признаков55 или выделять локальные варианты, опираясь па морфологическое своеобразие отдельных находок, в ряде случаев, действительно, резко выраженное, но не подтвержденное па групповом уровне56. В первом случае игнорируется имевшая место в верхнем палеолите и неотвратимая при сколько-нибудь широком расселении популяционная дифференциация, во втором — наблюдениям над характером индивидуальной изменчивости без достаточных оснований придается групповое значение. В результате в обоих случаях весьма вероятное действительное типологическое разнообразие верхнепалеолитического человечества остается невскрытым даже в географических рамках Европы, не говоря уже об африканских и азиатских популяциях, представленных единичными находками из районов, отстоящих друг от друга на тысячи километров. Способ выявления этих реальных типологических комбинаций признаков состоит, очевидно, в непредвзятом комбинировании наблюдений над индивидуальной изменчивостью отдельных находок и групповых характеристик, полученных для находок, сделаннйх в одном местонахождении пли близко одна от другой, в корректировании морфологического критерия различий с географическим и в сравнении выделенных вариантов с локальными расами в составе современного человечества. Какие признаки являются общими если и не для всех, то для большинства европейских находок верхнепалеолитического времени? Это большие и очень большие размеры горизонтальных диаметров черепной коробки, высота черепной коробки почти соответствует ее ширине, лобная кость довольно широкая, наклон се соответствует современным средним, ширина затылочной кости большая, лицевой скелет средней высоты (широко распространившееся даже в специальной литературе мнение о якобы очень низком лицевом скелете верхнеиалеолитических людей не подтверждается метрическими
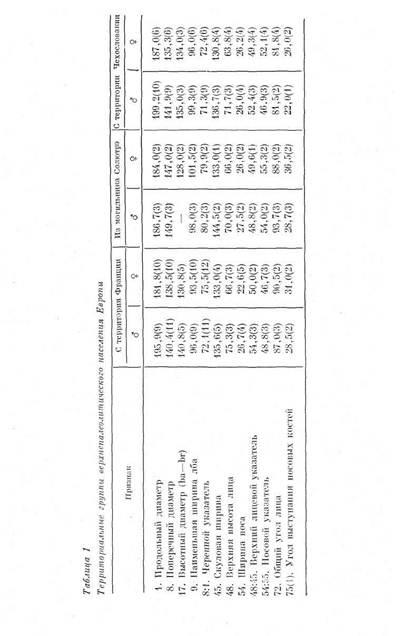
1 — женский неандертальский череп Гибралтар 1, 2 — мужской неандертальский череп Монте-Чирчео 1, 3 — мужской неандертальский череп Шапелль-о-Сен, 4 — женский неандертальский череп Штайнхайм, 5 — мужской неандертальский череп Брокен-Хилл, 6 — мужской неандертальский череп Шанидар 1, 7 — средняя по мужским черепам верхнепалеолитического времени из Европы, 8 — средняя по женским черепам верхнепалеолитического времени из Европы, 9 — минимум по современным монголоидам, 10 — максимум по современным монголоидам, 11 — минимум по современным европеоидам, 12 — максимум по современным европеоидам, 13 — черепа современных папуасов данными, высота лица соответствует средней европейской), нос широкий, орбиты чаще всего низкие, грушевидное отверстие среднеши-рокос, вертикальный профиль лицевого скелета ортогнатпый пли ме-зогнатный, горизонтальный профиль уплощенный в верхней части и достаточно острый в нижней, выступание носовых костей не уступает сильному выступаншо носа у европеоидов. Любопытно отметить, что индекс отношения нижнего угла горизонтального профиля к верхнему, столь эффективный, как мы помним, при отделении неандертальского вида от современного, оказывается пониженным на верхнепалеолитических черепах по сравнению с современными (рис. 6), демонстрируя, очевидно, следы не вполне закончившейся перестройки лицевого скелета в верхнепалеолитическое время. Таким образом, хотя комплекс перечисленных признаков в целом, как справедливо отмечал В. В. Бунак57, и не представлен в полной мере ни у одной из современных рас Европы, он достаточно характерен именно как европеоидный комплекс, стоящий у основания формирования европеоидной расы. Находки мужских черепов более многочисленны, поэтому они и положены в основу морфологического сопоставления. Нами уже отмечалось, что черепа из двух единственных верхнепалеолитических могильников, давших массовый материал, — Пшедмости и Солютрэ, вполне могут по своему морфологическому своеобразию претендовать на роль представителей самостоятельных вариантов 53. Черепа
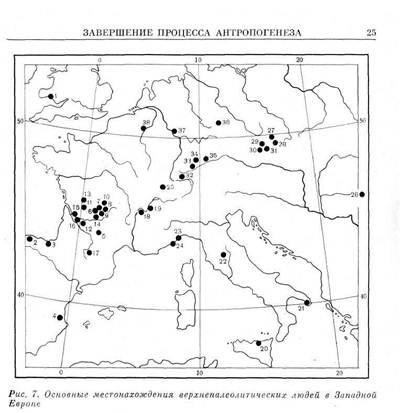
_____________ ЗАВЕРШЕНИЕ ПРОЦЕССА АНТРОПОГЕНЕЗА_____________ 23 из Пшедмости (Пшедмости I, III, IX) отличаются на фоне других находок тенденцией к усилению долихокрании, повышением и некоторым сужением лицевого скелета, широконосостыо. Черепа из Со-лютрэ (Солютрэ II, III, IV) резко брахикранны и очень широколицы, что может быть следствием прямой морфофизиологической зависимости, но более узконосы, чем черепа из Пшедмости. Подобные различия повторяются и на женских черепах. Таким образом, уже из сопоставления этих двух небольших серий вытекает, что верхнепалеолитическое население Европы не было единым по своему антропологическому составу и что внутри него выделялись местные варианты, отличавшиеся морфологическим своеобразием. Глядя на картину географического местоположения верхнепалеолитических находок (рис. 7), на которой представлены лишь те находки, которым можно дать метрическую характеристику, мы замечаем два территориальных «сгущения» верхнепалеолитических памятников, давших палеоантропологический материал: территорию Франции с долинами крупных французских рек и территорию Чехословакии. Чтобы увеличить численность сопоставляемых локальных групп и тем усилить статистическую значимость межгрупповых различий, вычислены средние по обеим территориям, опирающиеся на метрику основных находок. Для территории Чехословакии это, кроме Пшедмости, Дольни Вестонице, Брно, Младеч и. Павлов. Череп из Подбабы, ранее рассматривавшийся как верхнепалеолитический59, теперь должен быть определенно исключен из верхнепалеолитической серии, так как он имеет очень сомнительную датировку и, возможно, относится даже к раннему железному веку60. На территории Франции находки в Солютрэ, как указывалось, занимают самостоятельное положение, и поэтому в подсчет включены данные о черепах из всех остальных местонахождений, отличающихся более или менее сходной морфологией. Это следующие местонахождения: Кро-Маньон, Комб-Капель, Шанселяд, Ложори Басе, Рок де Сере, Ля Маделэн, Журдан, Вейрьер, Кап Бланк, Сен-Жермен-ля-Ривьер, Бидон, Лафайе, Плакар и Сорд. Средние по обеим группам - - французской и чехословацкой вместе со средними по черепам из Солютрэ представлены в табл. 1. Что демонстрирует нам эта таблица? Своеобразие солютрейского варианта очевидно. Его существование в верхнепалеолитическую эпоху свидетельствует о том, что брахикефализация — изменение формы черепной коробки в ходе времени, процесс, столь характерный для населения Европы в более поздние эпохи, — началась очень рано; фрагментарные данные по нижие-и среднепалеолитическим гоминидампозволяют датировать его начало эпохой нижнего палеолита и утверждать, что морфологические особенности солютрэйского варианта не являются случайными. Менее демонстративны отличия западноевропейских форм (французские находки) от центральноев-ропейских (Пшедмости, Брно и другие местонахождения Чехословакии). Центральноевропейский (по старой терминологии брюнн-
1 — Павилэнд, 2 — Камарго, 3 — Уртьяга, 4, — Парпайо, 5 — Бруникель, в — Лошери Басе, 7 — Кро-Маньон, 8 — Пато, 9 — Кап Блан, 10 — Ля Рогает, и — Шанселяд, 12 — Комб-Капелль, 13 — Плакар, 14 — Ля Шод, is — Сен-Жермен-ля-Ривьер, 1в — Рок де Сере, 17— Мае д'Азиль, is —Хото, 19 — Бейрьер, 20 — Сан-Теодоро, 21 — Романелли, 22 — Ольмо, 23 — Арен Кандид, 24 — Гримальди, 25 — Бишон, 26 — Чекловина, 27 — Младеч, 88 — Пшедмости, 29 — Брно, 30 — Дольни Вестонице, 31 — Павлов, 32 — Рётхе-корф, 33 — Штеттен, 34 — Кауферсберг, 35 — Нойессинг, 36 — Дёбриц, 37 — Оберкассель, 38 — Энгис пшедыостскпй, а, вернее сказать, брно-пшедмостский) вариант, по-видимому, действительно более длинноголов, чем западноевропейский (по старой терминологии кроманьонский). В высоте черепной коробки различия неопределенны, но лобная кость шире в централь-ноевропейской группе. В противовес старым взглядам, основанным на сравнении отдельных единичных черепов, нельзя утверждать, что центральноевропейская популяция более высоколица и узколица, чем западноевропейская: на мужских черепах вьтсоколицесть западноевропейского варианта видна отчетливо, хотя женские черепа
26 Глава первая
В старой палеоантропологической литературе под гримальдий- ским типом или вариантом подразумевалась негроидная комбинация признаков, которой якобы обладали два скелета из грота Детей в Гримальди — скелет пожилой женщины и юноши 15 — 16 лет 63. Однако, как уже упоминалось в начале этого раздела, подобная трактовка была возможна только вследствие того, что черепа были неправильно склеены из фрагментов, что усилило прогнатизм, птнроко-носость и уменьшило выступание носовых костей, т. е. придало им именно те признаки, на основании которых оказалось возможным говорить об их негроидном типе. Э. Влчек, осуществивший повторную и гораздо более тщательную реконструкцию, показал, что ни о каких негроидных признаках не приходится говорить в данном случае и что речь должна идти о вариациях признаков, более или менее типичных для верхнепалеолитического населения Европы в целом64. Он произвел и повторное измерение обоих черепов и любезно передал их автору этих строк (табл. 2). К сожалению, он не измерил углов лицевого профиля и выступания носовых костей, но на приложенных к его статье рисунках видно, что оба черепа стали много ортогнатнее, а носовые кости выступают па них значительно сильнее, чем раньше. Э. Влчек справедливо сомневается в правильности определения пола юношеского скелета и полагает, что его с
_____________ ЗАВЕРШЕНИЕ ПРОЦЕССА АНТРОПОГЕНЕЗА_______________ 27 Таблица 2 Черепа из грота Детей (Гримальди), традиционно относимые к «негроидному» типу
Признак 2, maturus 2, 15-16 лет Предыдущего черепа
8. Поперечный диаметр 131 132 133 9. Наименьшая ширина лба 98 96 98 48. Верхняя высота лица 59? 64? 69 54. Ширина носа 26 24 26 48: 45. Верхний лицевой указатель 46, 1? 54: 55. Носовой указатель 57, 8 50, 0 51, 8
В общем оба черепа из грота Детей, ранее считавшиеся негроидными, могут рассматриваться как черепа женских особей гримальдий-ской популяции в нашем понимании этого наименования. Мужской череп из того же грота Детей имеет «кроманьонский» облик и не отличается от мужских черепов из грота Барма Гранде, вместе они и образуют тот материал, морфологические особенности которого позволили нам выделить четвертый гримальдийский вариант.
|
 временному человеку. Далее важны открытые В. И. Кочетковой следы продолжающихся эволюционных преобразовании в мозгу верхнепалеолитических гоминид из Павлова и Кро-Маньона (череп Кро-Маньон III) 29, что говорит об отсутствии резко выраженной грани между характером эволюционного процесса на предсапиентной стадии и на стадии верхнепалеолитического человечества.
временному человеку. Далее важны открытые В. И. Кочетковой следы продолжающихся эволюционных преобразовании в мозгу верхнепалеолитических гоминид из Павлова и Кро-Маньона (череп Кро-Маньон III) 29, что говорит об отсутствии резко выраженной грани между характером эволюционного процесса на предсапиентной стадии и на стадии верхнепалеолитического человечества. При огромном разнообразии в строении черепа обезьян все же можно отметить, что относительно крупные формы (в том числе все человекообразные обезьяны, кроме резко специализированного гиббона) имеют значительный рельеф черепа — разнообразные гребни, служащие контрфорсами для прикрепления мышц, поддерживающих массивную нижнюю челюсть и способствующих поддержанию головы в определенном положении 35. Аналогичные костные образования на черепах ископаемых гоминид, начиная с древнейших, носят атавистический характер и редуцировались в ходе эволюции гоминид по мере редукции нижней челюсти и приобретения полностью выпрямленного положения тела. Разумеется, их постепенное исчезновение, замена гребней валиками, а затем исчезновение валиков — саггитального и надглазничного — не есть следствие «неупражнения» соответствующих образований, а результат, нужно полагать, отрицательной селекции, при которой обладание ненужными морфологическими структурами на черепе было обременительно в локомоторных актах, могло нарушать биомеханическую координацию движений и отсеивалось селекцией. Налицо, следовательно, тоже общий процесс, характерный для эволюции всех гоминид, но именно на стадии перехода от неандертальского человека к современному достигший апогея и приведший к исчезновению всех следов морфологических образований типа контрфорсов на покровных костях черепа современного человека.
При огромном разнообразии в строении черепа обезьян все же можно отметить, что относительно крупные формы (в том числе все человекообразные обезьяны, кроме резко специализированного гиббона) имеют значительный рельеф черепа — разнообразные гребни, служащие контрфорсами для прикрепления мышц, поддерживающих массивную нижнюю челюсть и способствующих поддержанию головы в определенном положении 35. Аналогичные костные образования на черепах ископаемых гоминид, начиная с древнейших, носят атавистический характер и редуцировались в ходе эволюции гоминид по мере редукции нижней челюсти и приобретения полностью выпрямленного положения тела. Разумеется, их постепенное исчезновение, замена гребней валиками, а затем исчезновение валиков — саггитального и надглазничного — не есть следствие «неупражнения» соответствующих образований, а результат, нужно полагать, отрицательной селекции, при которой обладание ненужными морфологическими структурами на черепе было обременительно в локомоторных актах, могло нарушать биомеханическую координацию движений и отсеивалось селекцией. Налицо, следовательно, тоже общий процесс, характерный для эволюции всех гоминид, но именно на стадии перехода от неандертальского человека к современному достигший апогея и приведший к исчезновению всех следов морфологических образований типа контрфорсов на покровных костях черепа современного человека. дертальскому виду может считаться решенным в положительном смысле, этим выбиваются морфологические аргументы из-под гипотезы пресаииенса, а сама гипотеза переводится в архив палеоантро-пологической науки. Таким образом, вопрос: существовал ли человек современного вида длительное время параллельно с неандертальцем, получает отрицательное решение.
дертальскому виду может считаться решенным в положительном смысле, этим выбиваются морфологические аргументы из-под гипотезы пресаииенса, а сама гипотеза переводится в архив палеоантро-пологической науки. Таким образом, вопрос: существовал ли человек современного вида длительное время параллельно с неандертальцем, получает отрицательное решение.
 мало различаются между собой в этом признаке. Вполне заметны отличия централышевропейского варианта от западноевропейского в вертикальном профиле лицевого скелета: цептралыюевропейские черепа прогнатнее, чем западноевропейские, — и вероятны в ширине носа: центральноевроиейская популяция как будто чуть более широконоса, хотя это и нельзя утверждать с большой определенностью. Если считать выделенные варианты реальными единицами попу-ляционной или надпопуляционной дифференциации верхнепалеолитического населения Европы, то встает вопрос о том, где проходила граница между ними. С этой точки зрения большое значение приобретают антропологические особенности скелетов из гротов Гримальди близ Ментоны61. Происходящие оттуда черепа крайней степенью долихокрагши, широколобостью и относительной шпрокоиосостыо напоминают центральноевропейские популяции, но они имеют орто-гнатный лицевой скелет, как и черепа западноевропейского варианта. Кроме этих особенностей, резко бросаются в глаза своеобразные особенности, присущие именно данной популяции, оставившей захоронения в гротах Гримальди: большие размеры черепа, огромная его высота, очень большая ширина и малая высота лицевого скелета, очень сильное выступание Носовых костей. Налицо комбинация признаков, которая напоминает «кроманьонский» тип в том понимании, которое вкладывали в этот термин старые авторы начала века 62. Так как эта комбинация несводима к рассмотренным выше, ее целесообразно выделить в самостоятельный четвертый вариант в составе верхнепалеолитического населения Европы и присвоить ему наименование гримальдийского.
мало различаются между собой в этом признаке. Вполне заметны отличия централышевропейского варианта от западноевропейского в вертикальном профиле лицевого скелета: цептралыюевропейские черепа прогнатнее, чем западноевропейские, — и вероятны в ширине носа: центральноевроиейская популяция как будто чуть более широконоса, хотя это и нельзя утверждать с большой определенностью. Если считать выделенные варианты реальными единицами попу-ляционной или надпопуляционной дифференциации верхнепалеолитического населения Европы, то встает вопрос о том, где проходила граница между ними. С этой точки зрения большое значение приобретают антропологические особенности скелетов из гротов Гримальди близ Ментоны61. Происходящие оттуда черепа крайней степенью долихокрагши, широколобостью и относительной шпрокоиосостыо напоминают центральноевропейские популяции, но они имеют орто-гнатный лицевой скелет, как и черепа западноевропейского варианта. Кроме этих особенностей, резко бросаются в глаза своеобразные особенности, присущие именно данной популяции, оставившей захоронения в гротах Гримальди: большие размеры черепа, огромная его высота, очень большая ширина и малая высота лицевого скелета, очень сильное выступание Носовых костей. Налицо комбинация признаков, которая напоминает «кроманьонский» тип в том понимании, которое вкладывали в этот термин старые авторы начала века 62. Так как эта комбинация несводима к рассмотренным выше, ее целесообразно выделить в самостоятельный четвертый вариант в составе верхнепалеолитического населения Европы и присвоить ему наименование гримальдийского. «Взрослые»
«Взрослые» 1. Продольный диаметр 191 192 198
1. Продольный диаметр 191 192 198 одинаковой вероятностью можно считать и мужским, и женским. Судя по размерам черепа, он принадлежал скорее женщине. На основании данных о динамике размеров черепа у человека современного вида восстановлены «взрослые» размеры юношеского черепа, подобно тому как это уже делалось во многих предыдущих случаях65.
одинаковой вероятностью можно считать и мужским, и женским. Судя по размерам черепа, он принадлежал скорее женщине. На основании данных о динамике размеров черепа у человека современного вида восстановлены «взрослые» размеры юношеского черепа, подобно тому как это уже делалось во многих предыдущих случаях65.


