ЛЕКЦІЯ № 9 2 страница
На постсинаптичній мембрані локалізовані хеморецептори, з якими може зв'язуватися медіатор, споріднений з ними, а це призводить до відкривання іонних каналів мембрани.
Нейротрансмітери
У ЦНС функцію трансмітерів виконує велика кількість біологічно активних речовин (близько ЗО). Належність синапсів до збудливих чи гальмівних визначається, з одного боку, специфікою медіаторів, аз іншого -видом рецепторів, які розташовані на постсинаптичній мембрані. Звичайно один і той же медіатор має на ній декілька рецепторів. Тому під час взаємодії одного і того ж трансмітера з різними рецепторами може виникати різний ефект - активація або гальмування постсинаптичного нейрона. Характер рецептора можна визначити не тільки за відмінністю ефекту, але й шляхом застосування хімічно активних речовин, що блокують передачу нервового імпульса через синапс (результат зв'язування з рецептором) або посилюють вплив трансмітера. Ці речовини можуть бути як ендогенного походження (утворюватися у самій ЦНС чи в інших органах і надходити до ЦНС через кров та лімфу), так і екзогенного (надходити із зовнішнього середовища) -сполуки, що надходять з їжею, лікарські препарати, різні отрути, токсини. Трансмітери ЦНС залежно від хімічної будови можна розділити на чотири основні групи: 1. Аміни (ацетилхолін, норадреналін, адреналін, дофамін, серотонін). 2. Амінокислоти (гліцин, глутамін, аспарагінова кислота, гамааміно- 3. Пуринові нуклеотиди. 4. Нейропептиди (гіпоталамічні ліберини і статини, оніоїдні пептиди, вазопресин, речовина Р, холецистокінін, гастрин тощо). У деяких синапсах виявлені також простагландини. У багатьох нейронах може міститися не один, а два чи більше трансмітерів. За своїм механізмом дії трансмітери можна поділити на два типи: іоно-тропні і метаботропні. Іонотропні трансмітери після взаємодії з рецепторами постсинаптичної мембрани змінюють проникність іонних каналів. На відміну від них метаботропні медіатори постсинаптичний вплив реалізують шляхом активації специфічних ферментів. У результаті в мембрані, а найчастіше - в цитозолі клітини активуються вторинні посередники (месенджери), які, у свою чергу, запускають каскади ферментативних реакцій у клітині або мембрані. Ці метаболічні зміни триваліші та глибші, ніж у разі дії іонотропних медіаторів. Вони можуть торкатися геному клітини, навіть передаватися спадково. Метаботропною активністю володіють більшість нейропептидів та низка інших трансмітерів, наприклад аміни. У разі виділення разом з основним медіатором вони здатні модулювати (підсилювати чи послаблювати) його ефект або регулювати його вихід. У результаті складних взаємодій, що відбуваються у ЦНС між трансмітерами і рецепторами, різними трансмітерами, ефект дії медіаторів може проявлятися по-різному. Так, ацетилхолін в одному випадку може викликати збудження, а в іншому - гальмування. Аналогічні ефекти характерні і для амінів. Серед амінокислот можна виділити ГАМК, що реалізує як пре-, так і постсинаптичне гальмування, гліцин, який реалізує постси-наптичне гальмування, та глутамінову кислоту, що передає збудження.
Механізм функціонування збуджувальних хімічних синапсів
Під впливом потенціалу дії відбувається деполяризація пресинаптич-ної мембрани, що зумовлює відкривання розташованих тут кальцієвих каналів. Іони кальцію пасивно (за градієнтом концентрації) надходять у пре-синаптичне закінчення, що сприяє наближенню міхурців з трансмітером до мембрани і, як наслідок, виділення його у синаптичну щілину. Трансмітер, дифундуючи через синаптичну щілину, досягає постсинаптичної мембрани, де з'єднується з відповідним рецептором, у результаті чого відкриваються хемозбудливі канали і підвищується проникність мембрани для іонів або Са2+. Це зумовлює деполяризацію мембрани -виникнення внутрішнього струму, що не досягає критичного рівня деполяризації. Такий потенціал називається збуджувальним постсинаптинним потенціалом (ЗПСП). Величина утвореного внаслідок цього струму є настільки малою, що він не здатний поширюватися, доки подібні впливи не деполяризують усю мембрану. ЗПСП, зумовлені активністю в основному синаптичному контакті, невеликі, проте деполяризації, що виникають у кожному з активних синапсів, додаються. Генерація ПД відбувається у результаті сумації ЗПСП. Цьому сприяють характеристики ЗПСП: порівняно велика його тривалість (наростання деполяризації 1-2 мс і спад - 10-12 мс), здатність поширюватися на сусідні ділянки мембрани, хоча і на невелику відстань. У постсинаптичній мембрані є не лише хемозбудливі, але й електрозбудливі канали. Тому перехід локального постсинаптичного потенціалу в потенціал дії відбувається у самій постсинаптичній мембрані. Сумація. Одним із варіантів сумації ЗПСП, що призводить до виникнення ПД, є часова сумація. Вона зумовлена тривалістю існування ЗПСП на постсинаптичній мембрані і виникає у разі дії повторних аферентних імпульсів в одному синапсі. Якщо до пресинаптичної мембрани за короткий проміжок часу будуть надходити ПД, то ЗПСП, що розвивається після кожного з них, накладається на попередній і в разі досягнення критичного рівня перейде у ПД. Таке явище можливе у зв'язку з тим, що звичайно по нервовому волокну надходять не поодинокі ПД, а їх групи - патерни ("пачки"). Інший різновид сумації - просторова. Якщо активність виникає одночасно в більше ніж одному синаптичному контакті, то простежується просторова сумація, й активність одного синапсу підсилює активність іншого, наближаючись до критичного рівня деполяризації. Синаптична затримка. У зв'язку із названим вище механізмом передачі збудження через синапс швидкість проведення у ньому сповільнюється, що зумовлено часом виділення трансмітера з міхурців пресинаптичної мембрани, часом дифузії його через пресинаптичну мембрану у синаптичну щілину, часом його дії на рецептори постсинаптичної мембрани, що називають си-наптичною затримкою. Синаптична затримка в ЦНС складає 0,2-0,5 мс.
5.3.3.1. Гальмівні синапси
Крім синапсів, що передають збудження, у ЦНС є і гальмівні синапси. Розрізняють два основні механізми гальмування: пресинаптичне і постси-наптичне. Пресинаптичне гальмування триваліше, ніж постсинаптичне. Його морфологічною основою є аксо-аксональні синапси, тобто аксон проміжного гальмівного нейрона утворює додатковий синапс на аксоні збуджувального нейрона. Гальмування розвивається завдяки зменшенню або блокаді виділення трансмітера в синаптичну щілину збуджувального синапса, тобто гальмування передачі нервових імпульсів відбувається завдяки зміні властивостей його пресинаптичної мембрани. Гальмування не поширюється на постсинаптичну мембрану такого синапса. Конкретний механізм пресинаптичного гальмування з'ясований недостатньо. Вважають, що трансмітер, який вивільняється в аксоні гальмівного нейрона, спричиняює стійку, тривалу деполяризацію закінчень збуджувальних нейронів. Протягом цього часу розвивається інактивація натрієвих каналів, унаслідок чого знижується збудливість мембрани аксона збуджуваль-
4 5-462 ного нейрона в ділянці гальмівного синапса. Тому потенціали дії або не доходять до збуджувального синапса, або їх амплітуда знижується і кількість вивільненого збуджувального трансмітера стає недостатньою для генерації ПД у нейроні, де відбувається процес гальмування. Можливий також інший механізм. Гальмівний трансмітер може зменшувати проникність пресинаптичної мембрани збуджувального синапса щодо іонів кальцію. Це призводить до зменшення викиду трансмітера в синаптичну щілину цього синапса, амплітуда ЗПСП у нейроні зменшується і він гальмується. Основним трансмітером для цього виду гальмування є ГАМК, а пригнічують його такі антагоністи, як бікукулін, пікротоксин (судомні отрути). У разі їх надходження в організм розвиваються судоми, оскільки велика кількість таких синапсів локалізована на мотонейронах спинного мозку. Постсинаптичне гальмування - це основний вид гальмування у ЦНС. Типовим гальмівним синапсом є аксо-соматичний синапс. Гальмівні синапси на тілі нейрона розташовані між збуджувальними си-напсами та аксонним горбком. Основними трансмітерами, що спричиняють цей вид гальмування, є ГАМК та гліцин. Кожен стимул, що прийшов до гальмівного синапса, викликає не деполяризацію, а, навпаки, гіперполяризацію постсинаптичної мембрани, що називається гальмівним постсинаптичним потенціалом (ГПСП). За своїм часовим розвитком ГПСП являє собою дзеркальне відображення ЗПСП із часом наростання 1-2 мс і часом спаду 10-12 мс. Підґрунтям гіперполяризації є підвищення проникності мембрани для іонів К+ і для іонів СІ. Відкриття хлорних каналів під впливом відповідних трансмітерів призводить до пасивного транспорту іонів хлору всередину нейрона, що гальмується, а надходження додаткових негативних іонів спричиняє збільшення від'ємного заряду мембрани. Оскільки гальмівний трансмітер може активувати калієві канали, то позитивно заряджені іони калію пасивно виходять з нейрона в інших синапсах, що також зумовлює її гіперполяризацію. Розвиток ГПСП перешкоджає досягненню нейроном критичного рівня деполяризації під впливом збуджувальних імпульсів, тобто знижує його збудливість. Таким чином, "боротьба" між ЗПСП і ГПСП визначає кінцевий результат: досягне деполяризація критичного рівня чи ні, виникне у нейроні ПД чи у відповідь на аферентні імпульси нейрон не проявить імпульсної активності й загальмується. Постсинаптичне гальмування значно поширене в ЦНС. Воно реалізується у нервових центрах, у мотонейронах спинного мозку, в симпатичних гангліях. У гангліях автономної нервової системи ГПСП реєструються протягом 30-100 мс, тобто більш тривало, ніж у соматичній нервовій системі. Унаслідок блокади даного виду гальмування розвиваються судоми (в разі надходження до організму стрихніну і правцевого токсину). Ме ханізми впливу стрихніну і правцевого токсину різні. Так, вважають, що стрихнін конкурує з гальмівним трансмітером на синаптичній мембрані, а правцевий токсин порушує його виділення з пресинаптичного закінчення. Відмінності цих двох видів гальмування полягають у тому, що пресинаптичне гальмування вибірково вимикає окремі входи до нервової клітини, у той час як постсинаптичне гальмування знижує збудливість нейрона в цілому; пресинаптичне гальмування триваліше, ніж постсинаптичне. Таким чином, локальні процеси гальмування, незважаючи на те, що не здатні до поширення, блокують проходження збудження, обмежують його поширення, запобігаючи цим перезбудженню ЦНС.
5.3.3.2. Втома нейрона і синапса
Більшість трансмітерів синтезуються у тілі клітини, накопичуються у вигляді гранул, а потім шляхом аксонного транспорту надходять до пресинаптичного потовщення, однак крім цього основного шляху трансмітер може синтезуватися і в самому нервовому закінченні. Особливо це характерно для холінергічних синапсів, де трансмітером є ацетилхолін. Однак коли через один і той самий синапс відбувається передача нервового сигналу протягом тривалого часу, то вміст трансмітера може зменшуватися. Це призведе до ослаблення швидкості передачі чи навіть повного її припинення. Такий стан називається втомою синапса. Швидкому розвитку втоми запобігають не тільки названі механізми відновлення вмісту трансмітера, але й функція астроцитів, спрямована на його ресинтез і передачу нервовому закінченню, що функціонує.
5.3.3.3. Електричні синапси
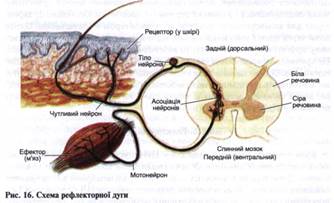
Крім типових хімічних синапсів у ЦНС зустрічається невелика кількість електричних синапсів. У них синаптична щілина значно вужча -близько 2 нм, тоді як у хімічних - до 20 нм. Опір мембрани в електричних синапсах низький, тому нервовий імпульс у вигляді ПД передається безпосередньо від пре- до постсинаптичної мембрани швидко, без синаптич-ної затримки, як і по нервовому волокну. Очевидно, що для електричних синапсів характерна двобічна провідність, мала стомлюваність. Різновидом електричної передачі ПД є ефаптпична, що здійснюється шляхом впливу позаклітинних струмів на сусідні клітини. І хоч у зв'язку з високим опором мембрани трансмембранні струми малі, вони можуть незначно змінювати величину мембранного потенціалу. Однак у деяких випадках (у разі ушкодження, хвороби) ймовірність ефаптичної передачі зростає. Можливо, що у зв'язку зі збільшенням частки електричної передачі нервових імпульсів у хворих за наявності певних порушень психічних функцій циркуляція нервових імпульсів може підтримуватися тривалий час, що порушує нормальну функцію ЦНС. 5.4. Основи рефлекторної діяльності центральної нервової системи Взаємодія різних нейронів ЦНС забезпечує її функціонування, підґрунтям якої є рефлекторна діяльність. Рефлексом називається стереотипна реакція організму або його окремих органів на сенсорний стимул, що реалізується за участі різних відділів нервової системи. Основою рефлекторних реакцій є рефлекторна дуга -комплекс утворів нервової системи, взаємодія яких забезпечує конкретний рефлекторний акт (рис. 16). Будь-яка рефлекторна дуга починається з рецепторів. Анатомічна ділянка тіла, що містить рецептори, під час подразнення яких виникає певний рефлекс, називається рецептивним полем рефлексу (рефлексогенною зоною). Звідси збудження поширюється по аферентних, вставних та еферентних нейронах до робочого органа.
Найменша рефлекторна дуга містить мінімальну кількість нейронів (два - аферентний та еферентний). Між ними є один синапс, тому ця дуга називається моносинаптичною. Більшість рефлексів - полісинаптичні. Час, що пройшов від початку дії подразника до появи відповідної реакції, називається часом рефлексу. Його тривалість залежить переважно від кількості синапсів, через які проходить нервовий імпульс, тому що швидкість поширення ПД по нервовому волокну набагато перевищує швидкість переходу його через синапс. Час рефлексу крім синаптичної затримки визначається й інтенсивністю стимулу, оскільки від цього залежить швидкість виникнення збудження у рецепторі і його частота. Класифікація рефлексів. За біологічним значенням рефлекси поділяються на харчові, судинні, рухові тощо. Залежно від того, де розташовані рецептори, рефлекси можуть бути екстероцептпивні (розташовані на зовнішній поверхні тіла), інтероцептивні (розташовані у внутрішніх органах) і пропріоцептивні (розташовані в опорно-руховому апараті - м'язах, сухожилках, суглобах). Залежно від локалізації нервового центру, в якому замикається рефлекторна дуга даного рефлексу, вони бувають спінальні (рефлекси спинного мозку), бульварні (рефлекси довгастого мозку), мезен-цефальні (рефлекси середнього мозку), діенцефальні (рефлекси проміжного мозку) і кіркові (рефлекси кори великого мозку). За характером відповідної реакції рефлекси поділяються на рухові (моторні), секреторні та судинорухові. За тривалістю рефлекторної реакції рухові рефлекси можуть бути фазичними (швидкими) і тонічними (тривала підтримка тонусу органа). За характером утворення і прояву рефлекси поділяються на безумовні (вроджені) й умовні (набуті у процесі індивідуального життя).
5.4.1. Фізіологічна характеристика нервових центрів Сукупність нейронів, що беруть участь у рефлекторній регуляції, об'єднуються у нервовий центр даного рефлексу. Це складне динамічне функціональне об'єднання, яке складається з багатьох "простих" центрів відповідальних за виконання найпростіших функцій. Динамічність цього утвору полягає у тому, що кожного разу задіяні саме ті структури ЦНС, які необхідні для виконання конкретного складного рефлекторного акту. Між ними може і не бути такого постійного зв'язку, який характерний для рефлекторної дуги. Він установлюється лише на час виконання якої-не-будь складної функції, що забезпечується наявністю широкого взаємозв'язку між нейронами ЦНС і властивостями нервових центрів. Нейрони, що утворюють нервовий центр, розташовані у визначеному порядку. Наприклад, кожна ділянка поверхні шкіри передає свої сигнали у певні ділянки спинного, проміжного мозку та кори великого мозку. Відповідне узгодження у нервовій системі цих сигналів дозволяє людині скласти уявлення як про тримірну структуру власного тіла, так і про його розташування у просторі. Така організація зв'язків у ЦНС, за якої просторове розташування нейронів, пов'язаних із певними рецепторами або виконавчими органами, що відображають специфіку їхньої просторової локалізації в організмі, називається соматотопічним представництвом функцій (зотаііс - тілесний, іорісиз - місцевий). Одночасно одна і та ж ділянка тіла іннервується значною кількістю нейронів - так звані зони перекриття (дублювання).
5.4.2. Властивості нервових центрів Процеси обробки інформації, що надходить до нервового центру, або формування команд до виконавчих органів зумовлені взаємодією нейронів між собою за допомогою синаптичних контактів. Дивергенція - це здатність нейрона встановлювати численні зв'язки з іншими нейронами. Завдяки цьому та сама клітина може брати участь у різних нервових процесах і реакціях, контролювати велика кількість інших нейронів, тобто кожен нейрон може забезпечити значне поширення імпульсів - іррадіацію збудження. Процеси дивергенції типовіші для аферентних відділів ЦНС. Конвергенція - це сходження різних шляхів проведення нейронних імпульсів до однієї і тієї ж нервової клітини. Конвергенція характерна для нервових центрів еферентних відділів. Нервові центри найчастіше представлені скупченням різних нейронів. Серед них зустрічаються як збуджувальні, так і гальмівні нейрони, нейрони сенсорні й моторні (аферентні чи еферентні). У нормі під час здійснення функції їх взаємодія дуже складна. Можна виділити такі властивості нервових центрів: 1. Сповільнення проведення збудження. Оскільки збудження (ПД) 2. Засвоєння і трансформування ритму. У нейронах ЦНС розрізня- Крім того, пейсмекерні клітини, маючи нижчий вихідний потенціал (до - 60 мв), легше збуджуються під час надходження імпульсів від інших відділів ЦНС. Імпульсні розряди нейронів, що виникають у відповідь на зовнішній подразник у пейсмекерних та інших нейронах, називаються викликаною активністю. У такому разі відповідь нервової клітини може виникати у вигляді поодинокого ПД або (найчастіше) серії імпульсів певної частоти. У разі ритмічних подразнень викликана активність нейрона може накладатися на ритм імпульсів, що надходять, і відбувається засвоєння ритму. Однак викликана активність нейрона нерідко не відповідає ритмові імпульсації, яка надходить. Нервові клітини мають властивість змінювати частоту імпульсів, що передаються, тобто мають властивість трансфор муванпя ритму. Вона зумовлена як наявністю у даному центрі різної кількості гальмівних та збуджувальних синапсів, так і властивостями подразника й функціональним станом клітини. За наявності високої збудливості нейрона сильний подразник може викликати таке зростання імпульсації клітини, що навіть на одиноке подразнення остання відповість залпом ПД.
5.4.3. Взаємодія рефлексів (координація рефлекторних функцій ЦНС) Регулювання багатьох складних функцій організму для забезпечення рефлекторної відповіді потребує участі кількох нервових центрів, що можуть бути розташовані навіть на різних ієрархічних рівнях ЦНС. Поява вищого рівня регулювання призводить до виникнення у ньому нових центрів регулювання, але із збереженням властивих їм функцій. У такому разі втрачається абсолютна автономність різних відділів ЦНС, усе більша частина функцій "передається" вищим відділам нервової системи. Взаємодія різних нервових центрів (як і взаємодія різних рефлексів) відбувається на основі визначених закономірностей. Вони дозволяють ЦНС здійснювати свої функціональні завдання щодо цілеспрямованого регулювання різних систем організму, його взаємодії із зовнішнім середовищем. Можна виділити такі основні принципи координації функцій ЦНС: 1. Важливою частиною нейронних ланцюгів, які утворюють рефлекторні дуги, є наявність гальмівних нейронів. Завдяки їм послаблюється або зовсім припиняється інтенсивний процес збудження, а також багато в чому впорядковується прояв рефлексу. Прикладом є реципрокне гальму-вання м'язів-антагоністів мотонейронів спинного мозку. Якщо відбуваються розлади центру, який регулює скорочення м'язів-згиначів, то центр м'язів-розгиначів одночасно загальмовується (і навпаки). Це здійснюється завдяки тому, що відросток аферентного нейрона, який несе збудження до нейронів-згиначів, розгалужується і його гілочка активує гальмівний нейрон. Аксон цього нейрона утворює гальмівні синапси на нейронах центрів розгиначів, що гальмуються за механізмом постсинаптичного гальмування. Другий досить поширений тип гальмування - зворотне гальмування, добре вивчене в спинному мозку, але характерне і для інших нервових структур. Аксони рухових нейронів спинного мозку, що передають збудження до відповідних скелетних м'язів, через свої колатералі спрямовують його також і до гальмівних нейронів (клітин Реншоу). Ці нейрони утворюють гальмівні синапси як на самих рухових нейронах (зворотне гальмування), так і на сусідніх {латеральне гальмування). У результаті досягається обмеження збудження мотонейронів (пригнічується їх надмірна імпульсація) та перезбудження (обмежується іррадіація збудження уздовж спинного мозку). Тому в разі введення в організм великих доз стрихніну, який блокує постсинаптичне гальмування, виникають тривалі виражені м'язові скорочення (судоми), що охоплюють усі м'язи скелета. За такої умови може настати смерть унаслідок судом дихальних м'язів і зупинки дихання. 2. Іррадіація і концентрація нервових процесів. Збудження, що вини- 3.Явища сумації й оклюзії. Сумація (накладання) виникає у разі дії кількох підпорогових стимулів (з різних рецепторів), кожний з яких, діючи окремо, не викликає відповіді. Просторова сумація їх сприяє посиленню відповіді нервового центра. Протилежне явище - оклюзія - розвивається за одночасної дії кількох надпорогових подразників, коли синаптичні поля розташовані поряд. У такому разі сумарна відповідь може бути меншою, ніж арифметична сума відповідей на кожен із подразників окремо. 4.Принцип "загального кінцевого шляху". Його підґрунтям є явище конвергенції. У ЦНС аферентних нейронів значно більше, ніж еферентних, тому на одному і тому ж еферентному нейроні збираються імпульси від кількох аферентних, тобто йде боротьба за загальний кінцевий нейрон. Отже, один і той же рефлекс можна викликати, подразнюючи різні рецептивні поля. 5. Домінанта. Зміст принципу полягає у тому, що за наявності одно-
5.5. Хімічні речовини та нервова система Певні хімічні речовини є необхідними для належного функціонування нервової системи. Серед них нейротрансмітери та нейромодулятори. у Хімічні речовини, що не є складовими нервової системи, можуть використовуватися для зміни (порушення) її нормального функціонування або для корекції порушених функцій. Ці хімічні речовини, чи ліки, поділені на кілька основних категорій (табл. 1). Стимулятори підвищують активність ЦНС, особливо головного мозку.
Серед них - амфетамін, кофеїн, кокаїн та нікотин. Ефективність їх впливу -від помірного (кофеїн) до сильного (метедрин). Депресанти пригнічують активність ЦНС, особливо головного мозку. До цієї групи належать: анестетики, барбітурати, алкоголь, опіати й транквілізатори. У помірних дозах барбітурати викликають сон, алкоголь знімає напруження, опіати полегшують біль, транквілізатори пригнічують страх (емоційне напруження). Однак у вищих дозах усі депресанти можуть завдати шкоди й призвести до розвитку патологічних реакцій, зумовити залежність. Антидепресанти послаблюють симптоми психологічної депресії. Поширене застосування трициклічних антидепресантів, таких, як тофраніл, що блокують нагромадження норепінефрину та серотоніну в нервових закінченнях. Психоделінні та галюциногенні засоби змінюють сприйняття і настрій. Негативні впливи включають галюцинації чи зміну сприйняття або навіть психози чи серйозні психічні розлади. Анальгетики послаблюють біль. їх поділяють на ненаркотичні, чи опіати, та наркотичні засоби. Опіати одержують з натуральних речовин, а наркотичні препарати є синтетичними. Обидві групи мають подібні фармакологічні властивості, включаючи здатність тамувати біль і, ймовірно, залежність від тривалості застосування. До анальгетиків відносять аспірин, кодеїн, меперидину гідрохлорид (демерол) і метадону гідрохлорид (долофін). Морфін є опіатом, що застосовується для боротьби із сильним болем, страхом та тривогою щодо болю. Транквілізатори застосовують для пригнічення страху та для сприяння сну. Майже всі вони широко використовуються. У разі застосування лише цих засобів вони порівняно нешкідливі, однак якщо зловживати ними, розвивається звикання. У разі поєднання їх з депресантами ЦНС чи алкоголем вони можуть бути небезпечними. До цієї групи відносять бензодіазегйни - діазепам (валіум), алпразолам (ксанакс), хлордіазепоксид (лібріум). Барбітурати також застосовуються як транквілізатори, але їх основна дія - снодійна До транквілізаторів виробляється звикання, а перевищення дози може бути фатальним. АВТОНОМНА НЕРВОВА СИСТЕМА
8.1. Функціональне значення автономної нервової системи
Французький фізіолог М. Біш (1801 р.) запропонував усі функції організму умовно поділити на соматичні (анімальні, або тваринні) і вегетативні (вісцеральні, рослинні). До соматичних функцій віднесені ті, що забезпечують взаємодію організму із навколишнім середовищем: сприйняття зовнішніх подразників і рухові реакції, що здійснюються скелетними м'язами. Автономними називають ті, від яких залежить здійснення обміну речовин у цілісному організмі, травлення, кровообіг, дихання, виділення, ріст і розмноження тощо. Відповідно до такого поділу функцій розрізняються і системи їх регулювання. Соматична нервова система забезпечує екстероцептивні сенсорні і моторні функції організму, автономна - регулює діяльність не тільки внутрішніх органів, але й автономні функції скелетних м'язів та власне нервової системи. Згідно із сучасною міжнародною анатомічною номенклатурою, терміном "автономна" замінено усі раніше існуючі: рослинна, вісцеральна, вегетативна. Поняття "автономна нервова система"включає комплекс центральних і периферійних нервових клітинних структур, що регулюють, забезпечують і контролюють функціональний рівень внутрішнього середовища організму для реалізації адекватних реакцій усіх систем на зміни чинників зовнішнього і внутрішнього середовища. Автономні компоненти реакцій організму довільно не контролюються. Саме тому автономну нервову систему і називають автономною, або мимовільною. Проте це поняття є дуже умовним, оскільки існують можливості умовнорефлекторного регулювання функцій внутрішніх органів, що вказує на участь кори великого мозку в регулюванні автономних функцій.
|