ЛЕКЦІЯ № 9 8 страница
Ністагм сприяє збереженню нормальної зорової орієнтації. Цю реакцію використовують також з діагностичною ме Будова і функції центральної нервової системи 313 тою для перевірки функціонування ста-токінетичиого аналізатора (присіпкового апарату). Площина ністагму збігається з площиною обертання, ось чому крім горизонтального розрізняють також вертикальний, діагональний та коловий ністагм. До статокіиетичиих рефлексів належать також рефлекси ліфта: ири підійманні в ліфті з прискоренням кінцівки розгинаються, а при опусканні — згинаються. Статичні і статокінетичпі рефлекси є у людини вже під час внутрішпьоутробіїого розвитку. Після народження, в міру дозрівання і вдосконалення центральних механізмів координації первово-м'язового апарату у дитини послідовно виникають рефлекторні реакції, які зумовлюють утримання голови, сидіння, стояння і ходіння. У подальшому відбувається вдосконалення процесів управління позами і рухами, зокрема розвивається кірковий контроль за функцією нервових центрів заднього мозку. 11.4.5. СЕРЕДНІЙ МОЗОК Середній мозок — це порівняно невелика частина стовбура головного мозку, розміщена між заднім (мостом) і проміжним мозком. У процесі еволюційного розвитку цей відділ мозку досить різко змінював свої розміри і функції. У вищих хребетних середній мозок поділяється на дві частини: покрівлю середнього мозку, розміщену над водопроводом середнього мозку (сільвієвим водопроводом), яка називається пластинкою покрівлі, або чотиригорбковою пластинкою, і ніжки мозку. Ніжки мозку заглиблюються у товщу середнього мозку; розділяються шаром сірої речовини, яка має значну кількість чорного пігменту меланіну і називається чорною речовиною, па дві частини — основу ніжок і покрив (мал. 160). Покрівля середнього мозку у риб і амфібій є вищим падсегмептарпим рівнем ЦНС, функціонально аналогічним передньому мозку вищих хребетних. У про
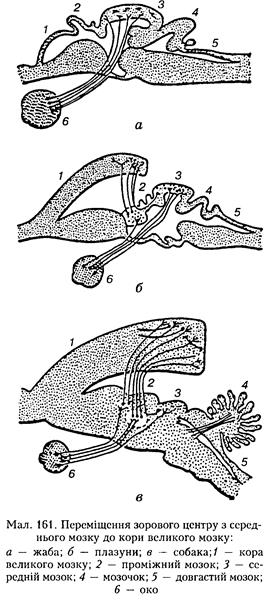
цесі ембріонального розвитку саме з цієї частини мозкової трубки формуються два мозкових пухирці, з яких пізніше утворюються очі. Тому у нижчих хребетних покрівля середнього мозку за своїми зв'язками і функціями є передусім вищим зоровим центром. Цей відділ середнього мозку у риб і амфібій (у них це двогорбко-ва пластинка) часто називають зоровими частками (покрівельною, тектальиою ділянкою). Покрівля середнього мозку має дуже складну мікроскопічну будову. Нейрони утворюють кірковий шар, а біла речовина розміщена всередині. Нервові елементи різних типів мають чітку орієнтацію, вони розміщені на різній глибині і тому утворюють 10 клітинних шарів. Основним аферентним шляхом є зоровий шлях, який прямує від сітківки до верхніх горбків (один із трьох підкіркових центрів зору). Другий потужний аферентний шлях прямує від ядра VIII пари черепних нервів до ядер моста, задніх горбків середнього мозку і колінчастих тіл метатала-муса. У риб цей нерв не має істотної слухової функції і головним чином проводить імпульси від присіикових рецепторів, а також від своєрідної аферентної системи — бічної лінії, яка викопує функції підтримання рівноваги тіла й орієнтації у ФІЗІОЛОГІЯ НЕРВОВОЇ СИСТЕМИ просторі і яка зникла після виходу хребетних па сушу. Крім того, кірковий шар покрівлі отримує значну кількість аферентних волокон від ядра трійчастого нерва. Отже, у нижчих хребетних він є вищим інтегративним центром усіх основних видів чутливості, крім нюхової. Проте провідною аферентацією цієї ділянки є зорова. Руйнування її у риб і земноводних робить тварин сліпими. Еферентні шляхи покрівлі середнього мозку прямують до моста і довгастого мозку і закінчуються переважно у їх сітчастому утворі. Частина аксонів покрівлі проходить через міст і довгастий мозок і прямує до спинного мозку, утворюючи покрівельно-снинпомозковий шлях. Прямі зв'язки структур покрівлі, зокрема ядра окорухового нерва, встановлено також з деякими ядрами моста і довгастого мозку. Після виходу хребетних на сушу функції покрівлі середнього мозку починають суттєво змінюватися. Це стосується насамперед зв'язків від ядер VIII пари нервів. Орган бічної лінії зникає, проте біля при-сіпкових ядер у задньому мозку виникають нові структури, пов'язані зі слуховою функцією, яка у наземних тварин досягає значного розвитку. У вищих рептилій і особливо птахів ця иейрониа структура дає початок заднім горбкам, і двогорбкова пластинка замінюється на чотиригорбкову. До задніх горбків надходять аферентні волокна від завиткових і частково при-сіикових ядер. Еферентні шляхи від нейронів цих горбків прямують до стовбурових ядер і сітчастого утвору. Передій горбки па цій стадії еволюційного розвитку все ще зберігають за собою функції вищого центру аналізу і координації зорової інформації. Проте у ссавців, яким властива значна кортикалізація функцій, виникають істотні зміни функцій чотиригорбкової пластинки. Якщо у рептилій і птахів аферентні шляхи, що йдуть від зорових і слухових рецепторів, закінчуються переважно у середньому мозку і лише незначна їх частина у вигляді колатералей потрапляє до проміжного мозку, де закінчується на ще не дуже розвинених колінчастих тілах, то у ссавців майже весь зоровий шлях переміщується вперед і закінчується у другому підкірковому і третьому центрах зору — бічному колінчастому тілі й подушці таламуса. І тільки невелика частина волокон зорового шляху відгалужується від них назад до передніх горбків чотиригорбкової пластинки. Отже, основний аналіз зорової інформації у ссавців переміщується до кінцевого мозку, а за середнім залишається лише деяка додаткова функція з обслуговування зорового апарату (мал. 161). Аналогічна ситуація спостерігається й зі слуховим аналізатором. Досліди із руйнуванням чотиригорбкової пластинки у ссавців підтверджують це положення: тварини з ушкодженою чоти-ригорбковою пластинкою не втрачають повністю зір і слухі, оскільки розрізняють світлові й звукові подразники, що переконливо доводиться методом умовних рефлексів. Проте деякі додаткові безумовні рефлекси, пов'язані з цими подразниками, у таких тварин зникають. Передні горбки чотиригорбкової пластинки середнього мозку у ссавців за допомогою окорухового нерва координують рухові реакції, пов'язані із зоровою функцією, тобто окорухові рефлекси, необхідні для нормального здійснення орієнтовних зорових реакцій та бінокулярного зору, а також низка вегетативних рефлексів, істотних для зору (скорочення внутрішніх м'язів ока, що забезпечують акомодацію ока і зіничний рефлекс). Задні горбки чотиригорбкової пластинки відповідають за рухові реакції, необхідні для нормального сприймання звуку і визначення його напрямку (підняття вух та їх повороти), а також такі, що пов'язані з появою звукових реакцій, які часто виникають у комплексі орієнтовної поведінки па раптові подразники. Так, під час подразнення задніх горбків дією раптового потужного звуку у тварин виникає мимовільний скрик. Усі ці орієнтовні реакції об'єднують під спільною назвою чотиригорбкового рефлексу. До нього входять також такі ком Будова і функції центральної нервової системи 315 іюнепти, як здригання і настороженість, а також низка складніших рухів, які формують реакцію уникнення. У ділянці покриву середнього мозку містяться червоне ядро, ядра деяких черепних нервів. Червоне ядро — це скупчення нейронів зі специфічною пігментацією. Воно відіграє важливу роль у регуляції автоматичних рухів, м'язового тонусу. Від нейронів червоного ядра починаються дві системи низхідних шляхів: червоноядерио-спиииомозковий шлях, що перехрещується після виходу з ядра, і червоиоядерио-оливні волокна, які прямують до нейронів сітчастого гігаитоклітиппого ядра, а також до переднього і заднього покривних ядер моста. Перетин стовбура мозку у кота уздовж переднього краю задніх горбків чотиригорбкової пластинки, тобто нижче від червоного ядра, призводить до виникнення децеребраційног ригідності, яка виявляється значним підвищенням тонусу переважно м'язів-розгиначів, тобто тих, що протидіють силам гравітації. Внаслідок цього відбувається сильне напруження і витягування кінцівок, спина вигинається, що голова і хвіст тварини підводяться догори (опістотонус). При цьому захисні згинальні рефлекси, які чітко виявляються у спіиальних тварин, різко загальмовані. Кінцівки можна зігнути, якщо прикласти значне зусилля, проте вони знову випрямляються, щойно це зусилля припиняється. Головною причиною децеребраційиої ригідності є блокада гальмівного впливу червоного ядра на бічне нрисіикове ядро (Дейтерса), розміщене у довгастому мозку. В нормі це ядро чинить збуджувальний вплив па рухові нейрони м'язів-розгиначів. Цей ефект значно підсилюється після зняття гальмівного виливу червоного ядра і рухових центрів, розміщених вище. Децеребраційна ригідність зникає після перетину мозку нижче бічного присіпкового ядра, а також задніх корінців спинного мозку. Децеребраційна ригідність не розвивається, якщо лінія перетину проходить
вище червоного ядра: в такому разі зберігається зв'язок середнього мозку з відділами ЦНС, розміщеними нижче. Отже, після децеребрації тварини значно підвищується збудливість рухових нейронів, що іннервують м'язи-розгипачі, і одночасно знижується тонус рухових нейронів м'язів-зги-иачів. 316 ФІЗІОЛОГІЯ НЕРВОВОЇ СИСТЕМИ У людини ригідність може виявлятися не лише внаслідок ураження нейроііиих структур середнього мозку, а й ири порушенні функції кори великого мозку і кірково-сіїишюмозкового шляху. Це пов'язано з тим, що у людини значно зростає роль кіркових структур у підтриманні м'язового тонусу і управлінні рухами тіла. Слід зауважити, що децеребраційна ригідність виявляється лише відносно анти-гравітаційних м'язів. Оскільки у людини в зв'язку з вертикальним положенням тіла силі гравітації протидіють па верхніх кінцівках м'язи-згипачі, то й ригідність у неї розвивається дещо інакше, ніж у більшості тварин: па верхніх кінцівках це виявляється у підвищенні тонусу м'язів-згиначів, а не розгиначів. Така сама картина і з тієї самої причини спостерігається у кажанів і лінивців. Центральна сіра речовина оточує водопровід середнього мозку. Саме тут ядра сітчастого утвору чинять полегшувальні впливи на кінцевий мозок, а також па деякі структури спинного мозку. У центральній сірій речовині локалізується антиноцицептивний (протибольовий) центр, збудження якого пригнічує больові відчуття. Крім згаданих структур у ділянці покриву середнього мозку розміщені парні ядра окорухового і блокового нервів, аксони яких іннервують зовнішні м'язи очного яблука, і одне непарне ядро (Едінгера) окорухового нерва парасимпатичні нейрони якого регулюють діаметр зіниці і здійснюють акомодацію ока. У ніжках середнього мозку розміщена чорна речовина. Вона складається з двох частий: сітчастої і щільної. Чорна речовина взаємодіє з білою кулею, хвостатим ядром, таламусом, червоним ядром, чотиригорбко-вою пластинкою, сітчастим утвором, корою великого мозку. Руйнування чорної речовини викликає значне зниження вмісту дофаміпу у хвостатому ядрі і є причиною порушень рухової функції, що виявляється у формі паркінсонізму. Вважають, що чорна речовина регулює додаткові співдружб рухи, що супроводжують основну рухову активність. 11.4.6. МОЗОЧОК Мозочок є типовою надсегментарною структурою, аферентні та еферентні зв'язки якої починаються і закінчуються в інших утворах мозку. Він формується па ранніх етапах філогенезу як елемент заднього мозку, який разом зі стовбуровою частиною цього відділу мозку з перших етанів свого існування здійснював регуляцію рухових функцій. У процесі еволюційного розвитку тваринного світу розміри мозочка і обсяг його функцій істотно змінювались. Оскільки основною функцією мозочка є координація рухів, то він добре розвинутий у риб і птахів, адже динаміка рухів у тримірному водному і повітряному середовищах потребує чіткої координації функцій м'язів. У амфібій і рептилій, які пересуваються по землі і мають, як правило, низько розміщений центр тяжіння, відбувається значна редукція мозочка. У зв'язку з вертикальним положенням тіла у людини розміри і значення мозочка істотно зростають. Досить добре розвинутий мозочок і в інших ссавців. Будова і зв'язки мозочка. Мозочок вищих тварин і людини складається з трьох відділів: правої та лівої півкуль, добре розвинених лише у ссавців, і розміщеного між ними черв'яка — давнього утвору мозочка. Структурпо-фупкціопально мозочок поділяють на три частки: клаптико-вузликову (ІоЬиз ііосиїопосіиіагіз) — стародавній мозочок (агспісегеЬеІІит); передню — давній мозочок (раїеосегеЬеІІит); задню — новий мозочок (пеосегеЬеІІит), що охоплює середню частину тіла і більшу частину півкуль мозочка (мал. 162). Зв'язок мозочка з іншими структурами мозку здійснюється за допомогою трьох пар ніжок. Нижні ніжки зв'язують мозочок зі спинним і довгастим мозком, най-товщі середні ніжки охоплюють довгастий мозок і, розширюючись, переходять на міст, що сполучає мозочок з корою великого мозку, а через верхні ніжки проходять еферентні та аферентні шляхи, зокрема передній спиниомозково-мозочко Будова і функції центральної нервової системи 317 вий шлях (Говерса), який є провідником проиріоцептивиої чутливості (від м'язів і суглобів). Аферентна імпульсація, яка потрапляє до мозочка, є специфічною для кожного з його відділів. Так, імпульсація до стародавнього мозочка (присінкомозочка) надходить нри-сіиково-мозочковими шляхами, які проходять через його нижні ніжки. Давній мозочок (спинномозкомозочок) отримує аферентну імпульсацію мохоподібними волокнами переважно від сиииномозко-во-мозочкових шляхів, основними з яких є задній (Флексига) і передній (Говерса) спиииомозково-мозочкові шляхи. Новий мозочок (мостомозочок) пов'язаний насамперед через міст з корою великого мозку і частково отримує інформацію також через спиииомозково-мозочкові шляхи. Сіра речовина — кора — мозочка має три шари: поверхневий молекулярний шар, шар грушоподібних нейронів (клітин Пур-кіньє) і нижній шар — зернистий. Крім кори в глибині мозочка серед білої речовини є скупчення нервових клітин, які утворюють його підкіркові парні ядра. Поверхня мозочка має велику кількість борозен, якими поділяється на окремі частки. У шарах кори мозочка розміщені шість різних тинів нейронів. Зерноподібні нейрони, кількість яких у людини становить 1010 - 10п, розміщені у зернистому шарі, а їхні аксони прямують до молекулярного шару, де розгалужуються Т-подібио, посилаючи в обох напрямках уздовж поверхні кори гілочки (паралельні нервові волокна) 1-2 мм завдовжки. Ці волокна проходять через ділянки розгалуження дендритів інших п'яти типів нейронів і утворюють на них синапси. У зернистому шарі розміщені також великі зірчасті нейрони (клітини Гольджі), дендрити яких поширюються на відносно невелику відстань, а аксони прямують до зерноподібних клітин (мал. 163). Шар грушоподібних нейронів налічує у людини до 15 мли нейронів, дендрити яких розгалужуються у молекулярному шарі. Аксони грушоподібних иейро
пів спускаються до ядер мозочка, а невелика їх кількість закінчується па при-сіпкових ядрах. Аксони останніх клітин становлять єдиний еферентний шлях від мозочка. Останні три типи нейронів — кошикові й зірчасті нейрони та клітини Лугаро лежать у молекулярному шарі, причому напрямок аксонів клітин Лугаро невідомий. Аксони кошикових нейронів закінчуються па тілі, а зірчастих — на дендритах грушоподібних нейронів. До кори мозочка входять два типи волокон. Висхідні нервові волокна (ліаио-иодібиі) проходять крізь зернистий шар і закінчуються у молекулярному на дендритах грушоподібних нейронів, а дуже численні (у людини — до 50 мли) мохоподібні волокна закінчуються на зірчастих нейронах. Кожне мохоподібне волокно віддає велику кількість колатералей, завдяки чому воно іииервує кілька нейронів. До кожного нейрона кори мозочка підходять численні паралельні нервові волокна від зірчастих клітин, і тому через ці нейрони на будь-якій клітині кори мозочка конвертують сотні мохоподібних волокон (див. мал. 163). 318 ФІЗІОЛОГІЯ НЕРВОВОЇ СИСТЕМИ
Майже половина соматосеисорпих шляхів, у тому числі спиииомозково-мозочкові шляхи (Говерса і Флексига) входять до мозочка у вигляді мохоподібних волокон. Решта — це спинномозково-оливиі волокна, котрі перемикаються в нижніх оливах на нейрони, що надсилають висхідні нервові (ліапонодібпі) волокна до кори мозочка. Деякі дослідники виділяють ще третю систему аферентних волокон — це адренергічні волокна, які надходять до кори мозочка від блакитного місця. Висхідні (ліапонодібпі) волокна утворюють численні збуджувальні сииапси на дендритах грушоподібних нейронів, і тому досить одного стимулу, щоб грушоподібний нейрон відповів цілим розрядом імпульсів. Мохоподібні волокна збуджують зірчасті нейрони, які через паралельні нервові волокна виявляють збуджувальний вилив на решту нейронів. Проте всі ці нейрони є гальмівними: великі зірчасті нейрони гальмують зірчасті нейрони, а розряди грушоподібних нейронів гальмують нейрони ядер мозочка. Отже, за винятком зірчастих, усі нейрони кори мозочка викопують гальмівні функції. Грушоподібним нейронам властива спонтанна імпульсна активність у стані спокою, чим і зумовлюється тонічне гальмування нейронів ядер мозочка. Зі зростанням активності цих гальмівних грушоподібних нейронів завдяки збудженню мохоподібних чи висхідних волокон гальмування нейронів ядер мозочка значно підсилюється. Якщо ж грушоподібні нейрони гальмуються (безпосередньо — зірчастими або кошиковими нейронами, опосередковано — за допомогою великих зірчастих нейронів), то це призводить до розгальмування структур мозочка, розміщених нижче. На думку Дж. Екклса (1969), велика кількість гальмівних нейронів у корі мо Будова і функції центральної нервової системи 319 зочка необхідна для припинення тривалої циркуляції імпульсів по нервових ланцюгах. Кожні 100 мс "обчислювальна машина" мозочка завдяки значним гальмівним зв'язкам скидає попередню інформацію і готова до приймання пової. Завдяки цій властивості мозочок може брати участь в оперативному управлінні рухом. Аксони грушоподібних нейронів закінчуються переважно па нейронах підкіркових мозочкових ядер (найбільше з них — зубчасте, ядро вершини (фастигіальие), коркоподібне й кулясте (інтерпозитиі). Нейрони від цих ядер прямують до стовбура головного мозку. Функціональне значення окремих ядер мозочка нерівнозначне. Так, коркоподібне й кулясте ядра відповідають за підтримання пози тіла, а зубчасте — за точність, швидкість і плавність рухів. Проте цілісна свідома і несвідома рухова функція здійснюється завдяки постійній взаємодії мозочкових і иозамо-зочкових структур. Функції мозочка. Мозочок впливає па функцію деяких автономних центрів, проте головна його роль — це забезпечення узгодженої рухової активності. Мозочок — головний керівний орган рухової системи, який здійснює координацію і контроль усіх видів рухів — від простих рухових актів до складних форм иоведіпкової рухової активності. Інформація про намір руху від асоціативних ділянок кори великого мозку через кірково-мозочкові шляхи (20 мли волокон) передається до півкуль мозочка і зубчастого ядра, де трансформується па програму дій, яка у вигляді нервових імпульсів повертається назад до рухових зон кори великого мозку переважно через бічні вентральні ядра таламуса. При здійсненні довільних рухів від рухової ділянки кори до певних рухових нейронів через кірково-спинномозковий шлях надходить велика кількість імпульсів, більше, ніж потрібно для виконання цієї рефлекторної реакції чи руху. Роль мозочка полягає у координації й корекції нервових імпульсів від кори великого мозку до виконавчих органів. Ця корекція відбувається так. Оскільки кожний довільний руховий акт здійснюється протягом певного часу, через кірково-спинномозковий шлях надходять еферентні імпульси до м'язів. За цей досить тривалий період передачі імпульсів від кори великого мозку до периферії і назад мозочок завдяки системі зворотного зв'язку з корою великого мозку встигає оцінити інформацію, що надходить від рухової зони кори, і забезпечує миттєву корекцію обсягу рефлекторних реакцій під час здійснення руху. Він вносить необхідні поправки у кількість імпульсів, що надходять від рухової зони кори великого мозку. Крім того, мозочок перешкоджає залученню до рухового акту "зайвих" груп м'язів. Однією з основних функцій мозочка у здійсненні рухових актів є також полегшення взаємодії антагоністичної мускулатури па початку і в кінці руху. Це забезпечує швидкий розвиток і закінчення рефлекторного акту. Наслідки уражень мозочка залежать насамперед від еволюційного рівня тварини. Так, якщо видалити мозочок у риб, це призводить до порушення функції рівноваги, риби не здатні плавати черевцем донизу, легко перевертаються на бік, па спину, хоча плавальні рухи у них не порушені. Це й зрозуміло, оскільки у риб мозочок є надбудовою над нрисінковим апаратом, який здійснює регуляцію положення тіла у просторі. Видалення мозочка у амфібій, рептилій і птахів спричинює складніші розлади рухової функції. У цих тварин також порушуються установчі рефлекси. Крім того, виникає погіршення регуляції тонусу скелетної мускулатури, переважно м'язів-розгиначів. Такі <(безмозочковіп тварини нагадують тварин з децеребраційною ригідністю, їхня рухова активність набуває некоординованого вигляду. Особливо значні порушення координації м'язової активності спостерігаються у птахів. У ссавців видалення мозочка крім зазначених симптомів порушення рівноваги і тонусу скелетної мускулатури призводить до розладнання регуляції довільних рухів. 320 ФІЗІОЛОГІЯ НЕРВОВОЇ СИСТЕМИ Руйнування невеликих частин мозочка не має значного впливу на здійснення рефлекторних реакцій завдяки компенсації функцій неушкоджеиими ділянками мозочка. Проте видалення половини мозочка призводить до тяжких порушень, які виникають па оперованому боці тіла: тварина набуває пози з витягнутими кінцівками і хребтом, викривленим у бік ураження, а голова при цьому повертається в здоровий бік. При спробі вставати тварина надає у бік ураження мозочка, а ири намаганні рухатись виникають манежні рухи, виявляється ністагм у контралатераль-пий бік. І хоча поступово порушення рухової активності зменшується, кінцівка па ушкодженому боці не може здійснювати нормального рухового акту, швидко втомлюється. При порушеннях функції мозочка у людини виникає розладнання координації рухів і м'язового тонусу. Типовим проявом уражень мозочка є тріада Шарко: ністагм, інтенційне дрижання (тремор) (виникає під час руху) і скандована мова, коли хворий не здатний координувати діяльність м'язів мовного апарату. Ушкодження мозочка добре компенсуються з боку інших структур ЦНС. Однак поступове руйнування мозочка внаслідок будь-якого патологічного процесу супроводжується сильним головним болем і запамороченнями, потім Порушується стояння і ходіння (атаксія). Деформована атаксична хода нагадує ходу п'яного, хворий широко розставляє йоги, ходить зигзагами, заточується, його "кидає" в обидва боки, іноді він надає навзнак. При пальце-носовій пробі хворий лише за третім разом влучає в ціль. Це зумовлено розвитком двох симптомів: асинергії, розладнання програми цілеспрямованих рухів, і дис-метрії, втрати співрозмірпості рухів та їх чіткості. Розрізняють також інші симптоми порушення функції мозочка у людини. Так, атонія виявляється значним ослабленням м'язового тонусу, який супроводжується симптомом астенії — швидкою стомлюваністю і внаслідок цього зниженням сили м'язових скорочень. Іноді такий хворий неспроможний встати з місця. Астазія (інтенційне дрижання) — це наявність переривчастих рухів через брак корегувального впливу мозочка. Лдіадохокінез — уповільнення реакції під час зміни одного типу руху на інший. При цьому хворі не можуть здійснювати швидку послідовність рухів, наприклад швидке згинання і розгинання пальців рук, оскільки внаслідок ураження мозочка не вмикаються антагоністичні м'язи при згашеній інерції в кіпці руху та на його початку — для подовження інерції спокою. У випадках ушкодження клаптико-вузлико-вої частки мозочка виникає дезеквілібра-ція — порушення рівноваги тіла. Отже, мозочок відіграє дуже важливу роль у регуляції пози та рухів. Якщо у тварин ураження чи видалення мозочка спричинює порушення тонусу мускулатури тіла і координації рухової активності, то у людини насамперед страждають тонкі й чіткі рухи (наприклад, писання, гра на роялі). Водночас у людей з такою природженою вадою, як відсутність мозочка, не спостерігається якихось істотних порушень рухових функцій. Це зумовлено тим, що мозочок — не єдиний орган, який регулює рухову функцію організму, і в разі його відсутності це здійснюють структури довгастого, середнього та кінцевого мозку. 11.4.7. АВТОНОМНА НЕРВОВА СИСТЕМА Автономна (вегетативна) нервова система (АНС) — це комплекс центральних і периферичних структур, які підтримують рівень гомеостазу, необхідний для адекватної реакції організму на впливи зовнішнього і внутрішнього середовищ. Автономна нервова система регулює ріст організму, його розмноження, обмін речовин, функції внутрішніх органів, забезпечує процеси адаптації організму до умов навколишнього середовища. Вона є функціонально автономною, тобто здійснює свої функції незалежно від нашої волі й свідомості. Будова і функції центральної нервової системи 321 Будова автономної нервової системи. Анатомічно і функціонально АНС поділяється на симпатичний та парасимпатичний відділи. Деякі вчені пропонують виділити метасимиатичиий відділ, який по суті є иіслявузловим відділом парасимпатичної нервової системи і охоплює комплекс вісцеральних вузлів, розміщених переважно інтрамуральио — у м'язовій стінці внутрішніх органів (серце, кишки тощо). Ця частина здійснює регуляторний вплив на функцію внутрішніх органів за допомогою місцевих периферичних рефлексів, що замикаються в її межах і можуть викопуватись за повної децентралізації органа (О. Д. Ноздрачов). Згадані вузли складаються з невеликої кількості нейронів (2-60), які синантично пов'язані між собою, утворюючи різноманітні сплетення. Характерною особливістю цих вузлів є їхня постійна фонова активність. До них підходять симпатичні й парасимпатичні нервові волокна, через які здійснюються центральні виливи. Особливістю будови автономної нервової системи є те, що еферентний шлях від ЦНС до органів-мішепей переривається у нервових вузлах (§ап§1іоп), утворених сукупністю останніх нейронів автономних рефлекторних дуг. Нервові волокна до автономних вузлів називають передвузло-вими (нрегаигліонариими), а ті, які виходять з вузлів, — післявузловими (ностган-гліонариими); вони прямують до виконавчих органів. Симпатична частина автономної нервової системи (симпатична нервова система) складається з двох відділів: центрального і периферичного. Центральний розміщений у бічних рогах сірої речовини спинного мозку (бічнопро-міжне ядро), починаючи від 8-го шийного до 3-го поперекового сегментів (мал. 164). Аксони нейронів цього ядра виходять із спинного мозку (иередвузлові волокна) у складі передніх корінців, відгалужуються від них і у вигляді білих сполучних гілок (мають мієлінову оболонку) підходять до симпатичних вузлів симпатичного стовбура. Вищі інтегративні, регу
ляториі й координаційні центри - див. с. 324, 328. Периферичний відділ складається з парного симпатичного стовбура і нервів, що відходять від нього, проміжних вузлів і сплетень. Із вузлів симпатичного стовбура виходять післявузлові волокна у вигляді сірих сполучних гілок (не мають мієлінової оболонки), які прямують на периферію до виконавчих органів. Парасимпатична частина автономної нервової системи (иарасимпа- 322 ФІЗІОЛОГІЯ НЕРВОВОЇ СИСТЕМИ
Мал. 165. Парасимпатична частина автономної нервової системи (передвузлові волокна зображено суцільними лініями, післявузлові — штриховими): / — війчастий вузол; післявузлові волокна до: 2 — зіниці; З — сльозової залози; 4 — привушної залози; 5 — піднижньо-щелепної залози; 6 — серця; 7 — бронхів; 8 — шлунка; 9 — кишок; 10 — печінки; // — підшлункової залози; 12 — нирок; 13 — товстої кииіки; 14 — сечового міхура; 15 — статевих органів. Латинськими цифрами позначено черепні нерви; 52-54 — сегменти спинного мозку тичиа нервова система) також має центральний і периферичний відділи. Центри знаходяться в середньому мозку, в дорсальній частині моста, у довгастому мозку, в сірій речовині 2-4-го крижових сегментів спинного мозку (мал. 165). Вищі інтегративні, регуляторні й координаційні центри - див. с. 328. Периферичний відділ — це передвузлові волокна, що йдуть у складі окорухового, лицевого, язикоглоткового і блукаючого нервів, вузли (війкові, крило-ніднебінні, вушні, під'язикові, иіднижньощелеині), тазові нутрощеві (вісцеральні) нерви, вузли та їхні зв'язки. Розрізняють дві частини парасимпатичної нервової системи: черепну і тазову.
|