ЛЕКЦІЯ № 9 12 страница
Під впливом інсуліну підвищується проникність клітинної мембрани і для амінокислот, з яких синтезуються білки. Внаслідок уведення великих доз інсуліну різко знижується рівень глюкози в крові. Насамперед відчувають цей дефіцит головний і сиишшй мозок, оскільки глюкоза є основним джерелом енергії для нервових клітин. Коли вміст глюкози знижується до 2,5 ммоль (45-50 мг%), виникає гостре порушення функції мозку — інсуліновий шок (кома). Вивести з такого етапу може внутрішньовенне введення розчину глюкози. Зниження вмісту інсуліну в крові внаслідок недостатньої функції підшлункової залози або експериментальної її екстирпації призводить до цукрового діабету, що супроводжується гіперглікемією, глюкозурією та іншими порушеннями. Головним стимулом для виділення інсуліну є підвищення концентрації глюкози в крові. Крім того, такий самий ефект спричинює подразнення блукаючого нерва. Подібність ефектів, що виникають під виливом інсуліну і блукаючого нерва, взаємозв'язок між ними зумовили об'єднання їх у єдину вагоінсулярну систему. Другий гормон підшлункової залози — глюкагон, фізіологічна дія його пов'язана насамперед з вуглеводним обміном. Він збільшує рівень глюкози в крові за рахунок розпаду глікогену в печійці і в цьому є синергістом адреналіну. На виділення глюкагону виливає зниження рівня глюкози в крові й соматотропін гіпофіза. Симпатична стимуляція підвищує секрецію глюкагону.
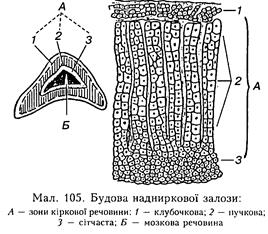
Надниркові занози 233 9.5. НАДНИРКОВІ ЗАЛОЗИ Надниркові залози парні, нагадують сплющені піраміди із заокругленою вершиною. Складаються з кіркової і мозкової речовин. Кіркова речовина має три зони: клубочкову (зовнішню), пучкову (середню) і сітчасту (внутрішню). Усі зони чітко відмежовані (мал. 105), і їх кіркові ендокриноцити виробляють різні гормони: клубочкова — міпералокортикоїди, пучкова — глюкокортикоїди, сітчаста — статеві гормони. У риб кора і мозковий шар надниркових залоз є парними міжиирковими незалежними органами. Починаючи з амфібій, намічається сполучення надниркових парних тілець у надниркові залози. У рептилій і птахів залози складаються з кіркової і мозкової речовий і представлені у вигляді смужки біля аорти. З кори надниркових залоз виділено понад 40 стероїдних сполук, однак лише вісім з них мають високу біологічну активність. Міпералокортикоїди (альдостерон, кортикостерон, дезоксикортикостероп) беруть участь у регуляції мінерального обміну в організмі, насамперед натрію і калію в плазмі крові. З цих гормонів найбільшу активність має альдостерон. Він збільшує реабсорбцію натрію в каиальцях нирок, що забезпечує підвищення його вмісту в крові, і разом з тим знижує реабсорбцію калію, що призводить до його втрати. Підвищення концентрації натрію в крові під виливом альдостерону призводить до затримки води в організмі і сприяє підвищенню артеріального тиску. Нестача міие-ралокортикоїдів призводить до втрати натрію, що спричинює зміни у внутрішньому середовищі організму, несумісні з життям. Через кілька днів після видалення кори надниркових залоз настає смерть. Врятувати життя може лише введення мінерало-кортикоїдів і натрію. Рівень мінералокор-тикоїдів у крові регулюється кількістю натрію і калію. Натрій гальмує секрецію альдостерону і виділяється з сечею. Має також значення співвідношення концентрацій йопів натрію і калію. Це підтверджує той факт, що підвищення секреції альдостерону зумовлює як дефіцит натрію, так і підвищення вмісту калію в крові. Усі ці регулювальні впливи здійснюються через гіпоталамус. Глюкокортикоїди (кортизон, гідрокортизон і кортикостерон) регулюють вуглеводний, білковий і ліпідний обмін. Найактивнішим з них є гідрокортизон. Глюкокортикоїди підвищують рівень глюкози в крові, але не за рахунок глікогену печінки, а шляхом перетворення безазотистих залишків дезаміповапих амінокислот па вуглеводи (глюконеогенез). Глюкокортикоїди не є життєво необхідними гормонами, і все ж їх дефіцит призводить до зниження опірності організму щодо шкідливих чинників. У етапі стресу активізується виділення адеиогіпофізом кортикотропіну, а під його впливом і глюкокортикоїдів у надниркових залозах, що підвищує захисні реакції організму. Глюкокортикоїди збільшують загальну кількість лейкоцитів при зменшенні кількості еозинофільних гранулоцитів. Ця реакція є настільки сталою, що отримала визнання в експериментальній роботі й у клініці для оцінки функціонального стану кори надниркових залоз під назвою проби Торна. Виділення глюкокортикоїдів регулюється за участю рилі-зипг-гормоиів гіпоталамуса, зокрема кор-тикотроиіи-рилізипг-гормоиу. На гіно-
234 ВНУТРІШНЯ СЕКРЕЦІЯ таламус впливає насамперед адреналін, який виділяється внаслідок рефлекторних впливів, під час болю, ири крововиливах, різких температурних виливах, інфекційних захворюваннях, психічних травмах тощо. Статеві гормони кори надниркових залоз (андрогени, естрогени) відіграють важливу роль у розвитку статевого апарату дітей, коли ще недосконала ендокринна функція статевих залоз. Підвищена секреція статевих гормонів кори надниркових залоз призводить у дітей до передчасного статевого дозрівання. Після настання статевої зрілості їх роль є незначною. Проте після припинення функції статевих залоз у старості кора надниркових залоз залишається єдиним джерелом андрогенів і естрогенів. Мозкова речовина надниркових залоз є складовою частиною симпато-иадиирко-вої (-адрепалової) системи, що забезпечує секрецію похідних катехолу катехол-амінів — адреналіну і норадрепаліну. Вони руйнуються в крові і тканинах ферментом аміноксидазою. Мозкову тканину надниркових залоз називають хромафінною (клітини забарвлюються солями хрому). Клітини, що продукують гормони в цій тканині (мозкові ендокриноцити), є видозміненими ніслявузловими симпатичними нейронами. Вони іннервуються иередвузловими волокнами симпатичної нервової системи. Адреналін і норадреналіи є симпатичними медіаторами, тому їхня дія подібна до дії симпатичної нервової системи. Адреналін підвищує систолічний артеріальний тиск, активізує роботу серця, підвищує рівень глюкози в крові за рахунок глікогену печійки, збільшує вміст вільних жирних кислот у плазмі крові, підвищує рівень основного обміну, знижує тонус гладких м'язів шлунка і кишок, пригнічує їх рухову активність, підвищує тонус скелетних м'язів. У високих дозах спричинює аритмію і екстрасистолію внаслідок підвищення збудливості серця. Незалежно від дії на серце адреналін звужує кровоносні судини, але сприяє розширенню стінки артеріол внутрішніх органів (шлунка, печінки, кишок). Норадреналін виливає па серцево-судинну систему своєрідно. Якщо адреналін зумовлює тахікардію, то норадреналін — брадикардію внаслідок виливу блукаючого нерва. При внутрішньовенному введенні адреналін підвищує активність нервової системи. Катехоламіни підтримують гомеостаз в умовах змін зовнішнього середовища, сприяють забезпеченню функціонуючих систем кров'ю, ЇЇ перерозподілу в загальній системі кровообігу. Таким чином вони мобілізують резерви організму па боротьбу зі шкідливими стимулами (стресорами) і залучають інші адаптивні механізми, що підвищують опірність організму. 9.6. СТАТЕВІ ЗАЛОЗИ Статеві залози є мішаними, оскільки разом із продукцією статевих клітин — спер-матозооиів і яйцеклітин — виділяють у кров статеві гормони — андрогени й естрогени. Обидві групи статевих гормонів утворюються як у чоловічих, так і в жіночих статевих залозах. Однак у чоловіків превалюють андрогени, а в жінок — естрогени. За хімічною природою статеві гормони є стероїдами. Ці гормони необхідні для статевого дозрівання, забезпечення розвитку вторинних статевих ознак (мал. 106) і виконання статевих функцій. Внутрішньосекреторну функцію сім'яників виконують іитерстиційні клітини (клітини Лейдіга). Це великі клітини, розміщені між сім'яними каиальцями біля кровоносних капілярів. Вони продукують андрогени — тестостерон, апдростероп та інші, найактивнішим серед яких є тестостерон. Тестостерон здійснює статеву диференціацію організму в ембріональний період. Під його впливом відбувається маскулінізація зовнішніх і внутрішніх статевих органів. Із настанням статевої зрілості андрогени стимулюють сперматогенез і розвиток вторинних статевих ознак, сприяють збільшенню гортані, потовщенню голосових зв'язок, що змінює тембр голосу. Андрогени Статеві занози 235
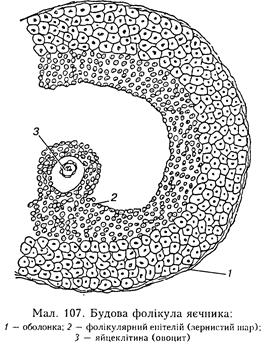
стимулюють синтез білка, що забезпечує розвиток мускулатури, виливають па функцію нервової системи і визначають статеву поведінку самців. Нестача статевих гормонів викликає зміни, які залежать від того, виникли вони задовго до статевого дозрівання чи після. У першому випадку статеве дозрівання припиняється і статеві органи не досягають зрілого стану, не розвиваються вторинні статеві ознаки. У другому випадку в статевій системі відбуваються лише часткові зміни. У регуляції функції чоловічих статевих залоз провідну роль відіграють гонадотропіни аденогіпофіза. їх уведення прискорює і посилює розвиток чоловічого статевого апарату і вторинних статевих ознак. Видалення гіпофіза у статевозрілих тварин призводить до атрофії сім'яних капальців та іитерстиційної тканини. Внутрішньосекреторна функція яєчників. Яєчник продукує естрогени (ес-традіол, естрон) і прогестини (прогестерон та ін.). Естрадіол виробляється у фолікулах яєчника, прогестерон є гормоном жовтого тіла. Продукція естрогенів пов'язана з фолікулярним епітелієм (зернистим шаром) третинних (пухирчастих) фолікулів (граафових пухирців) (мал. 107). Кількість жіночих статевих гормонів залежить від фаз статевого циклу. Так, естрогени створюють умови для запліднення яйцеклітини, а прогестерон забезпечує імплантацію і розвиток зародка після запліднення. У прямій залежності від функції яєчника перебуває матка. Після введення естрогенів відбувається гіпертрофія матки, ЇЇ набрякання, ріст маткових труб і посилення скоротливості їхніх м'язів, що сприяє переміщенню яйцеклітини до матки. Основним місцем утворення прогестерону є клітини жовтого тіла. Жовте тіло виникає в кожному статевому циклі після овуляції на місці зруйнованого фолікула. Другим джерелом прогестерону є плацента. Цей гормон називають також гормоном вагітності, оскільки він не тільки забезпечує нормальний розвиток заплідненої яйцеклітини, а й гальмує дозрівання чергового фолікула та овуляцію.
236 ВНУТРІШНЯ СЕКРЕЦІЯ З настанням статевої зрілості у жінок овуляція повторюється періодично через кожні 28 днів — менструальний цикл. Вій має чотири періоди: 1) иередовуляцій-пий; 2) овуляційиий; 3) иісляовуляційний; 4) період міжовуляційпого спокою. У тварин статевий цикл називають естральним. Він пов'язаний з тічкою і властивий всім ссавцям, крім мавп і людини. У передовуляційному періоді відбувається підготовка до імплантації яйцеклітини — до вагітності. Матка збільшується, епдометрій стає пухким, його залози розростаються, скоротливість маткових труб посилюється. Вміст гонадотронінів в аде-ногіиофізі збільшується як у першому, так і в другому періоді, а після овуляції знижується. Під впливом гонадотронінів посилюється вироблення яєчником естрогенів, які й зумовлюють зазначені зміни в статевому апараті, дозрівання фолікула й овуляцію. Настання овуляції забезпечує вихід яйцеклітини і переміщення її в матковій трубі. Після запліднення яйцеклітина потрапляє в матку і закріплюється в її слизовій оболонці. На місці фолікула розвивається жовте тіло, клітини якого продукують прогестерон. Під впливом прогестерону продукція гонадотронінів аденогіпофі-зом зменшується, що, у свою чергу, зменшує утворення яєчниками естрогенів. 9.7. ЗАГРУДНИННА Це центральний орган імуногенезу, який визначає становлення і функціонування клітинної системи імунітету. Залоза міститься за грудниною у верхньому відділі переднього середостіння (мал. 108) і складається з двох часток, у яких є дві групи клітин — тимоцити та лімфоцити. Єдиної думки про залозу як орган внутрішньої секреції немає. Проте пересадження
Якщо запліднення не відбулося, настає післяовуляційний період. Виникає менструація як результат скорочення матки і відторгнення її слизової оболонки, підготовленої до вагітності, що не відбулася. Після цього настає період міжовуляційпого спокою. Секреція естрадіолу і прогестерону регулюється гаподотропінами гіпофіза — фо-літропіном (фолікулостимулювальпим гормоном) і лютропіном (лютеїнізуючим гормоном). У перші дні статевого циклу наростає кількість фолітроніну, що стимулює дозрівання первинного фолікула до вторинного, далі — третинного. Принципове значення має співвідношення між фолі- і лютропіиом. У середині циклу зростає рівень лютроиіну, що спричинює розрив третинного фолікула і перетворення його на жовте тіло. Плацента продукує пролактин (лак-тогенний гормон) і релаксин, який до моменту пологів сприяє підвищенню податливості лобкового сполучення. Під час пологів підвищується рівень иейросекрету нейрогіпофіза окситоципу, який активізує скоротливість м'язів матки і сприяє скороченню міоепітеліальпих клітин в альвеолах молочних залоз, разом з пролактином забезпечуючи лактацію. На функцію статевих залоз значний вплив чинить ЦНС. Під впливом негативних емоцій перебіг статевого циклу може змінюватися. ЗАЛОЗА (ТИМУС)
Шишкоподібна заноза (епіфіз). Тканинні гормони залози після хірургічного видалення її відновлює властиві їй функції. Виділено кілька біологічно активних речовин залози: тимозии, тимопоетші та ін. Під впливом тимозину відбувається диференціація попередників Т-лімфоцитів до імупокомие-теитиих Т-лімфоцитів. Тому захворювання, що характеризуються імунними порушеннями, пов'язують із порушенням функції загрудниниої залози. Існує думка про участь залози в еритропоезі. Встановлено, що у випадках тимоми — пухлини залози — розвивається особливий вид анемії, коли з периферичної крові зникають ре-тикулоцити, а з кісткового мозку — еритробласти. Численні експериментальні дослідження свідчать про наявність зв'язків між загрудиинною і статевими залозами. Тривале введення високих доз екстрактів загрудниниої залози молодим тваринам спричинює затримку статевого дозрівання. 9.8. ШИШКОПОДІБНА ЗАЛОЗА (ЕПІФІЗ) Шишкоподібна залоза, або шишкоподібне тіло (епіфіз), у нижчих хребетних є фо-торецептивним органом і має назву тім'яного ока. У ссавців ця залоза розміщена в ділянці чотиригорбкової пластинки середнього мозку і функціонує як орган внутрішньої секреції. З неї виділено мелатонін, який зумовлює затримку статевого розвитку в статевонезрілих особин, а в дорослих самок гальмує статевий цикл. Крім того, мелатонін гальмує виділення лютроиіи-ри-лізииг-гормоиу, а звідси секрецію гонадотронінів і активність статевих залоз. Секреції мелатоніну властивий циркадний (добовий) ритм, максимальний рівень секреції спостерігається вночі. Світло гальмує секрецію мелатоніну. Гальмування секреції мелатоніну протягом світлового дня збільшує кількість ліотропіи-рилізииг-гормоиу і го-падотропіиів, викликає менструацію (тічку), ріст сім'яників, статеву активність. Екстирпація шишкоподібної залози спричинює гіперглікемію, а введення її екстракту — гіпоглікемію. У дослідах на плазунах показано, що мелатонін спричинює агрегацію гранул меланіну в меланоцитах шкіри, що зумовлює посвітління шкіри. Отже, він є антагоністом меланотроиіну (мелапоцитсти-мулювальпого гормону) проміжної частки гіпофіза. 9.9. ТКАНИННІ ГОРМОНИ Крім залоз внутрішньої секреції біологічно активні речовини продукуються спеціалізованими клітинами різних органів. Зокрема, це стосується підішжішощелеішої слинної залози, яка виділяє інсуліноподібний білок і калікреїн, що забезпечує судинорозширювальний ефект. Нирки виділяють ренін і еритропоетин. Клітини кишок синтезують цілу низку гормонів, які впливають на секрецію, моторику і всмоктування речовин: секретин, холецистокінін (панк-реозимін), гастроінтестинальний, вазоін-тестинальний пептиди (ГІП, ВІП), бомбе-зин, мотилін, вілікінін, соматостатин тощо. Шлунок продукує гастрин і гістамін. Нещодавно відкрито групу нейрорегу-ляторних пептидів — енкефалінів, ендор-фінів, нейротензину та ін., які становлять ще одну гормональну систему гіпотала-мо-гіиофізарного комплексу. Ці речовини мають гіпофізотроппу активність і споріднені з рилізииг-гормоиами. Частину з них внесено до АРІЮ-системи (Атіпе Ргесаг-зогз иріаке апй ОесагЬохуІаііпд-зузіет) — системи захоплення попередників амінів та їх декарбоксилувапня. Клітини цієї системи мають високий вміст амінів. їм також властива здатність виробляти крім пептидів біогенні аміни (серотонін, до-фамін, гістамін).
ГУМОРАЛЬНЕ РЕГУЛЮВАННЯ ФІЗІОЛОГІЧНИХ ФУНКЦІЙ
Механізми регуляції усіх життєвих функцій організму прийнято ділити на нервові та гуморальні. Перші використовують для передачі і переробки інформації структури нервової системи (нейрони, нервові волокна) та імпульси електричних потенціалів, другі - внутрішнє середовище організму й молекули хімічних речовин. Обидва механізми забезпечують єдину нейрогуморальну регуляцію функцій органів та їх систем. Гуморальна регуляція - різновид біологічної регуляції, являє собою засіб передачі інформації до ефекторів через рідке внутрішнє середовище організму (кров, лімфу і тканинну рідину) за допомогою молекул хімічних речовин, які виділяються клітинами або спеціалізованими тканинами й органами. Гуморальну регуляцію підрозділяють на місцеву, малоспеціалізовану саморегуляцію і високоспеціалізовану систему гормональної регуляції, яка забезпечує генералізовані ефекти за допомогою гормонів. Місцева гуморальна регуляція (тканинна саморегуляція) практично не керується нервовою системою, тоді як гормональна регуляція складає частину єдиної нейрогуморальної системи. Поділ механізмів регуляції життєдіяльності на нервові й гуморальні умовний, насправді нервові та гуморальні механізми регуляції нероздільні, тому необхідно говорити про єдину нейрогуморальну систему регуляції автономних функцій. Гормональна регуляція забезпечується ендокринною системою, до складу якої входять: 1) ендокринні органи або залози, основною і єдиною функцією яких є 2) ендокринна тканина в органі, тобто скупчення інкреторних клітин в 3) клітини органів, які володіють крім основної одночасно й ендокрин- Місцева регуляція забезпечується такими основними засобами, підґрунтям яких є передача хімічних сигналів у межах однієї тканини або органа за допомогою креаторних зв'язків клітин, простих метаболітів і більш складних продуктів обміну речовин - біологічно активних речовин, а також електролітів та медіаторів. 9.1. Поняття про гормони
Генералізовані спеціалізовані ефекти гормональної регуляції здійснюються за допомогою особливих хімічних регуляторів внутрішнього середовища - гормонів. Гормонами називають хімічні речовини, що утворюються і виділяються спеціалізованими ендокринними клітинами у внутрішнє середовище організму для регуляції обміну речовин та вегетативних функцій організму, гуморального забезпечення координації й інтеграції процесів життєдіяльності. Синтез гормонів ендокринними клітинами відбувається безперервно, його інтенсивність залежить не тільки від регуляторних сигналів ланок управління, але й від величини секреції. Гормони можуть депонуватися у тих же тканинах, де утворюються. Депонування гормонів ендокринною тканиною може здійснюватися у спеціалізованих гранулах (мозкова речовина надниркових залоз) або у спеціалізованих структурах залози (колоїд фолікулів щитоподібної залози). Гормони переносяться рідинами внутрішнього середовища як у вільній, так і в зв'язаній формі. Діють вони так: а) надходять до рідких середовищ організму і переносяться ними до віддалених клітин-мішеней (телекриновий ефект); б) місцево на сусідні клітини (паракриновий ефект)] в) на саму клітину (автокриновий ефект). Для регуляції функціонування багатьох органів та процесів цей механізм хоч і діє більш повільно, але виявляється ефективнішим, ніж нервова регуляція. Це зумовлено наступним: а) біологічно активна сполука може надходити до кожної клітини; б) спектр зазначених регуляторів значно ширший, ніж нейротрансмітерів; в) їх дія триваліша. Серед біологічно активних сполук виділяють: 1.Справжні гормони - секретуються залозами внутрішньої секреції (тироксин, інсулін, паратгормон тощо) або спеціалізованими ендокринними клітинами, що розташовані в різних органах (ліберини, ста-тини, гормони травного тракту тощо). 2.Тканинні гормони - секретуються неспеціалізованими клітинами різних органів (гістамін, серотонін, простагландини, енкефаліни тощо). 5. Метаболіти - речовини, що утворюються у результаті обмінних процесів у тканинах (молочна кислота, аденозин, вуглекислий газ, деякі іони тощо). Саме тому поняття "гуморальна регуляція" ширше ніж "гормональна регуляція", яка здійснюється лише за допомогою гормонів. Виділяють п'ять видів дії гормонів: і) метаболічну (вплив на обмін речовин); 2) морфогенетичну (стимуляція формотворення, диференціювання органів і тканин, росту); 3) кінетичну (включення певної діяльності); 4) коригувальну (зміна інтенсивності функцій органів і тканин); 5) реакто-генну (здатність змінювати реактивність тканин). За хімічною природою усі гормони поділяються на три групи: 1)похідні амінокислот - гормони щитоподібної залози, мозкового шару надниркових залоз, епіфізу; 2)пептидні гормони, прості і складні білки - гіпоталамічні нейропепти-ди, гормони гіпофізу, острівкового апарату підшлункової залози, прищиподібних залоз; 3)стероїдні гормони - гормони кіркового шару надниркових залоз, статевих залоз, гормон ниркового походження - кальцитріол. На основі функціональних критеріїв розрізняють три групи гормонів: 1) гормони, які впливають безпосередньо на орган-мішень; ці гормони називають ефекторними\ 2) гормони, основною функцією яких є регуляція синтезу і виділення ефекторних гормонів, їх називають тропними; 3) гормони, що синтезуються нервовими клітинами в гіпоталамусі, вони регулюють синтез і виділення гормонів аденогіпофізом, їх називають рилізинг-гормонами.
Загальні властивості гормонів 1.Специфічність дії. Кожен гормон діє на конкретні фізіологічні системи, органи чи тканини, тобто на ті структури, що містять спеціалізовані рецептори до нього. 2.Дистантна дія. Багато гормонів діють через внутрішнє середовище на органи, які розташовані далеко від місця їх утворення. Проте тканинні гормони, що секретуються без участі спеціалізованих ендокринних клітин, найчастіше проявляють місцеву дію на невеликій відстані від місця їх утворення. 3.Не мають видової специфічності, за винятком гормону росту і р- ліпотропіну. 4.Дія зумовлена впливом на клітини-мішені плазматичних мембран. 5.Висока біологічна активність - вплив на функції органів і тканин у дуже невеликих концентраціях. Транспорт гормонів кров'ю. Гормони циркулюють у крові у вільному стані або у вигляді сполук з форменими елементами крові. Більшість гормонів (близько 80%) переносяться у комплексі зі специфічними білками плазми крові, які вибірково транспортують молекули того чи іншого гормону Після сполучення з білками гормони переходять у неактивну форму, а також стають захищеними від руйнування ферментами крові. Концентрація більшості гормонів у крові непостійна. З одного боку, вона визначається інтенсивністю утворення і секреції гормону залозою, а з іншого - швидкістю його поглинання, руйнування та виведення. Час напіввиведення (Т 1/2) гормонів із крові становить понад 4 доби - у тироксину, кілька годин - у стероїдів, хвилини - в інсуліну, вазопресину, ка-техоламінів і секунди - у нейромедіаторів та пептидів. Секреція деяких гормонів є періодичною. Наприклад, рівень гормону росту, АКТГ, має виражені добові коливання, які пов'язані з циклами неспання. А для статевих гормонів характерна не лише добова ритміка утворення і секреції, але й чітко виражена періодика, пов'язана із віковими періодами розвитку, менструальним циклом та циклом "вагітність-пологи". Далеко не завжди в організмі людини гормони утворюються у кількості, що забезпечує їх нормальне функціонування. Функціональна активність залози може бути підвищеною (гіперфункція) або, навпаки, зниженою (гіпофункція). Ця зміна може бути короткочасною, що зумовлено необхідністю змінити активність відповідної функції організму, або мати більш тривалий чи постійний характер і торкатися змін не тільки функції, але й структури ендокринної залози, що характерно для стану хвороби.
Взаємодія гормонів
У кров надходить велика кількість гормонів із різних залоз внутрішньої секреції, кожен з яких може впливати на кілька функцій організму. Одна і та ж функція, один і той самий орган знаходяться під впливом кількох гормонів, які здійснюють сумарну фізіологічну дію. Цю взаємодію гормонів можна розділити на три види - синергізм, антагонізм і пермісивна дія. Синергізм. Часто кілька гормонів, що впливають на функцію органа, здійснюють односпрямовану (синергічну) дію. Наприклад, адреналін і глюкагон активують розпад глікогену в печінці до глюкози та зумовлюють збільшення рівня цукру в крові. Загалом, це прояв принципу дублювання, що забезпечує високу надійність регуляторних механізмів. Антагонізм гормональних впливів часто відносний. Так, інсулін та адреналін здійснюють на рівень глюкози в крові протилежний вплив: уведення інсуліну зумовлює гіпоглікемію, а адреналіну - гіперглікемію. Однак спільне біологічне значення ефекту цих гормонів проявляється у поліпшенні вуглеводного живлення тканин. Аналогічним чином проявляється взаємодія гормону росту й інсуліну. Діабет частіше виникає у повних людей. Це пояснюється ліполітичним ефектом соматотропного гормону, що підсилює резистентність клітин (особливо скелетних м'язів) до інсуліну. Виникає, з одного боку, підвищена потреба клітин у глюкозі, з іншого - підвищується вміст глюкози у плазмі крові. Одночасно сомато-тропний гормон пригнічує активність бета-клітин підшлункової залози, що продукують інсулін. На основі описаних фактів склалося уявлення про гормони та антигормони. Наприклад, гормон гіпофізу мелатонін гальмує розвиток статевих залоз і функцію щитоподібної залози. Антикортикотропін, що продукується в епіфізі, пригнічує синтез альдостерону наднирковими залозами. Прогестерон пригнічує синтез естрогенів. Можливо, у деяких випадках, наприклад, у разі зміни середовища у клітині (рН, концентрації іонів, температури) різною мірою виявляється дія того чи іншого гормону. Наприклад, замість ефекту меланоцитостимулювального гормону гіпофіза виявляється дія АКТГ кори надниркових залоз, подібного з ним за структурою, і навпаки. Пермісивна дія гормонів виявляється у тому, що сам гормон не викликає фізіологічного ефекту, але створює умови для реакції клітини чи органа на дію іншого гормону. Наприклад, глюкокортикоїди не впливають прямо на тонус м'язів судин і на розпад глікогену в печінці, однак вони створюють умови, за яких надграничні концентрації адреналіну збільшують артеріальний тиск і спричиняють гіперглікемію (за рахунок посилення глікогенолізу в печінці). Сформована система співвідношення гормонів визначає їх сумарний фізіологічний ефект на функціональний стан людини (пубертатний період, старість, вагітність тощо). Цей стан залежить від гормонального профілю, тобто кількості різних гормонів у крові людини. Власне кажучи, гормональний профіль - це рівень активності залоз внутрішньої секреції, які визначають активність різних гормонів у плазмі крові, функціонування тканин, що захоплюють гормони й утилізують їх, а також їх виведення з організму. Тому необгрунтоване застосування гормонів як лікувальних препаратів може спричинити небажані перебудови гормонального профілю усього організму.
Механізм дії гормонів
Дія гормону (ліганда) на клітину реалізується шляхом взаємодії з рецептором. Рецептори можуть бути розташовані як на мембрані, так і всередині клітини. Ті гормони, що погано проникають через мембрану (кате-холаміни, пептидні гормони), фіксуються на ній зовні. У такому разі їх вплив на функціональний стан клітин здійснюється за допомогою посередників. Стероїдні і деякі тиреоїдні гормони, що легко проникають у клітину, мають здатність впливати на внутрішньоклітинні структури, діють на процеси транскрипції, змінюючи активність синтезу клітинних білків. Однак нерідко і ці гормони виконують свої функції також за допомогою месеиджерів посередників (рис. 29). Стероїдні гормони. Стероїдний гормон легко проникає у клітину завдяки малим розмірам молекул і високій сприйнятливості ліпідів мембран. У результаті у визначених клітинах вміст гормону стає високим, хоча у крові він знаходиться у надзвичайно малих концентраціх. Після проникнення стероїду в клітину відбувається його зв'язування із внутрішньоклітинним рецептором і транспорт через цитоплазму та ядерну мембрану до ядра. Наступний етап - "доставка" гормону до тієї ділянки генома, у якій відбувається його зв'язування з кислим білком (гістоном) хромосоми. Зв'язування відбувається саме з тією ділянкою генома, яка забезпечує транскрипцію іРНК і наступний синтез специфічного білка в цитоплазмі клітини. Результатом такого впливу стероїдів є зміна синтезу білків у
|