ЛЕКЦІЯ № 9 7 страница
Слід зазначити, що рефлекс розтягання м'язів-розгиначів у людини відіграє дуже важливу роль у протидії силі гравітації і відповідно підтриманні вертикального положення тіла. Тому вій постійно перебуває під контролем вищих нервових центрів — гальмується ири довільному згинанні кінцівки або посилюється за допомогою механізму, що називається у-петлею, коли виникає потреба нерухомо чи струнко стояти тривалий час. Моносинагітичпий рефлекс розтягання у людини можна також спричинити електричною стимуляцією найбільш збудливих аферентних нервів групи Асе, що йдуть від м'язів. Рефлекторна реакція, що виникає при цьому, називається Н(ашУрефлексом. Цей рефлекс звичайно викликають електричною стимуляцією великогомілкового нерва під коліном, реєструючи електроміограму (ЕМГ) камбалоиодібиого м'яза зовнішніми електродами. При слабких стимулах (20-30 В) з'являється спочатку тільки короткочасна рефлекторна реакція у вигляді Н-хвилі ЕМГ з латентним періодом 30-35 мс (Н-рефлекс). Зі збільшенням сили подразнення виникає збудження й еферентних нервових волокон, які активують весь м'яз з латентним періодом 5-10 мс (М-рефлекс). Деякий час реєструються обидві рефлекторні реакції (Н- і М-реф-лекси), але при подальшому зростанні сили 304 ФІЗІОЛОГІЯ НЕРВОВОЇ СИСТЕМИ
подразнення на осцилоірамі можна помітити лише М-відповідь (мал. 154). Це зменшення Н-відиовіді при підвищенні сили стимуляції зумовлено антидромпими ПД в еферентних нервових волокнах, які спричинюють зворотне гальмування в рухових нейронах. Проте слід зазначити, що навіть найпростіший моиосииаитичний рефлекс у багатьох випадках виконується за участю не лише збудливих, а й гальмівних антагоністичних нейронів. Цей гальмівний зв'язок здійснюється через вставний нейрон, який, збуджуючись, виділяє гальмівний медіатор і за його допомогою викликає гальмування рухового нейрона, що відбувається, зокрема, при реципрокиому гальмуванні (див. с. 296, мал. 150). Полісинаптичні рефлекси — це такі реакції, центральна частина рефлекторної дуги яких складається з кількох нейронів. Більшість таких нейронів є вставними, а останній, аксон якого йде до ефектора, — еферентним. У випадку рухових полісинаитичних рефлексів таким останнім нейроном є руховий нейрон. Полісинаптичні рефлекси відіграють важливу роль у локомоції, харчовій і харчопошуковій функціях, захисті організму від шкідливих подразників. Одним з таких иолісинаптичиих рефлексів є перехресний згинально-розгинальний рефлекс, який забезпечує крокування наземних тварин і здійснюється па рівні спинного мозку. Цей рефлекс виявляється у тому, що на кінцівці, яка згинається і робить крок вперед, відбувається скорочення м'язів-згипачів, розслаблення м'язів-розги-начів, а па протилежній (коптралатераль-ній), яка в цей момент є опорною, — одночасно скорочення м'язів-розгиначів і розслаблення м'язів-згиначів. Наступний крок відбувається в результаті зміни фаз роботи м'язів-антагоиістів обох кінцівок па протилежні. Така взаємодія кінцівок під час крокування пояснюється тим, що під час збудження центру м'язів-згиначів однієї Будова і функції центральної нервової системи 305
кінцівки відбувається рецинрокие гальмування центру м'язів-розгипачів цієї самої кінцівки і одночасно збуджується центр м'язів-розгипачів і гальмується центр м'я-зів-згипачів контралатеральної кінцівки. Механізм такого гальмування дослідив Дж. Екклс. Так, аферентні нервові волокна, тобто довгі аксони біполярних нейронів чутливого вузла спинномозкового нерва, входячи у спинний мозок, розгалужуються, причому одна гілка збуджує рухові нейрони, що іннервують м'язи-згипачі, а інша — нейрони, які утворюють гальмівні синанси па рухових нейронах, що іннервують м'я-зи-розгииачі (звичайно за допомогою клітин Реишо). Отже, подразнення аферентного волокна викликає збудження центру м'язів-згииачів і гальмування центру м'язів-розгипачів іпсилатеральпої кінцівки і одночасно спричинює протилежні реакції у центрах м'язів контралатеральної кінцівки (мал. 155). Вісцеральні рефлекси. Крім рухових спинний мозок здійснює велику кількість вісцеральних рефлексів, за допомогою яких регулює функцію внутрішніх органів і систем. Так, через спинний мозок замикаються регіопарпі судинорухові рефлекси, у крижовому відділі знаходяться центри сечовиділення, дефекації і статевих функцій. Звичайно, ці рефлекси перебувають під контролем вищих відділів ЦНС, зокрема кори великого мозку, проте вони зберігаються і у спіпальпої тварини, хоча стають менш досконалими і неконтрольовапими. Вони реалізуються через автономну нервову систему і детальніше будуть розглянуті нижче. 11.4.2. ДОВГАСТИЙ МОЗОК Довгастий мозок — це невелика частина головного мозку, яка значною мірою зберегла ознаки будови ембріональної мозкової трубки. Сіра речовина довгастого мозку утворює скупчення — ядра черепних нервів. Ці ядра і нерви, що відходять від них, частково зберігають сегментарну будову довгастого мозку, властиву спинному мозку. Центральний канал спинного мозку в довгастому мозку перетворюється на четвертий (IV) шлуночок, дію якого утворює ромбоподібну ямку, яка є дорсальною поверхнею моста і довгастого мозку. Ядра черепних нервів. У довгастому мозку розміщені ядра V, VII —XII пар черепних нервів; (V — трійчастий; VII — лицевий; VIII — нрисінково-завитковий; IX — язикоглотковий; X — блукаючий; XI — додатковий; XII — під'язиковий) (мал. 156). Для кращого, цілісного сприймання розглянемо усі дванадцять пар черепних нервів, незважаючи па те, що вони належать до периферичної частини нервової системи. Нюховий (І) і зоровий (II) нерви — цс чутливі нерви. Перший іннервує нюхові рецептори нюхового епітелію носа. Зоровий нерв утворений шаром нервових волокон — аксонів мультиполяр- 306 ФІЗІОЛОГІЯ НЕРВОВОЇ СИСТЕМИ
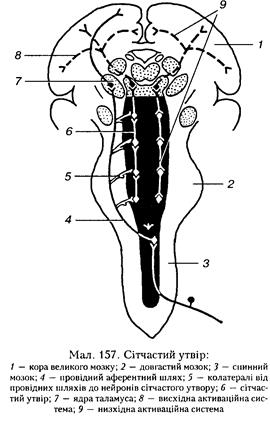
них нейронів гангліозного шару сітківки ока (див. мал. 175, с. 352). Окоруховий (III) і блоковий (IV) — рухові нерви, забезпечують координацію рухів очей, іннер-вуючи зовнішні м'язи очного яблука. У складі окорухового нерва є парасимпатичні волокна, що іннервують гладкі волокна війкового м'яза і м'яза — звужувача зіниці. Ці волокна беруть участь у здійсненні зіничного рефлексу. Трійчастий нерв (V) — змішаний, його рухові волокна іннервують жувальні м'язи і м'язи, що змінюють натяг барабанної перетинки. У разі однобічного ураження цих волокон чи нейронів нижня щелепа під час відкривання рота зміщується в бік ураження. Чутливі волокна проводять збудження від рецепторів шкіри всього лиця, слизової оболонки носа, кон'юнктиви, окістя черепних кісток, зубів, твердої мозкової оболонки і смакових рецепторів язика. Відвідний нерв (VI) іннервує прямий м'яз очного яблука. Його ураження викликає зниження тонусу цього м'яза, зіниця зміщується в напрямку до носа. Лицевий нерв (VII) — змішаний, його еферентні волокна іннервують всю мускулатуру лиця, а також сльозові, піднижньощелепні і під'язикові слинні залози. Через аферентні волокна лицевого нерва проводиться збудження від смакових рецепторів передньої частини язика. Присінково-завитковий нерв (VIII) є чутливим нервом. Він складається з двох гілок — завиткової і присінкової. Завиткова гілка проводить збудження від рецепторів спірального (кортієво-го) органа завитки внутрішнього вуха, а присінко-ва — від рецепторів півколових проток і мішечків присінка. Язикоглотковий нерв (IX) є змішаним: складається з рухових, чутливих і парасимпатичних волокон. Рухові волокна іннервують м'язи м'якого піднебіння, глотки, гортані і голосових зв'язок, беручи участь у здійсненні рефлексів чхання, кашлю і блювання. Парасимпатичні волокна іннервують привушні слинні залози. Чутливі волокна проводять збудження від рецепторів сонної пазухи (каротидного синуса), слухової (євстахієвої) труби і стінок барабанної порожнини, а також від смакових рецепторів задньої третини язика. Блукаючий нерв (X) є найбільшим нервом людського організму. Він також є змішаним — містить рухові, парасимпатичні й чутливі волокна. Чутливих волокон у складі блукаючого нерва близько 95 %. Вони передають інформацію від рецепторів ротової порожнини, глотки, значної частини травного каналу, дихальних шляхів, легень, рефлексогенних зон судин і серця, а також від твердої мозкової оболонки і вушної раковини. Рухові (парасимпатичні) волокна блукаючого нерва іннервують гладкі м'язи більшої частини травного каналу, трахеї, бронхів, серцевий м'яз, а також травні залози шлунка, тонкої кишки, підшлункову залозу, печінку, нирки. Додатковий нерв (XI) — руховий, іннервує груднино-ключично-соскоподібний і трапецієподібний м'язи. При порушенні функції цього нерва або його ядра знижується тонус цих м'язів і плече на боці ураження опускається, що утруднює чи навіть унеможливлює повертання голови у протилежний бік. Під'язиковий нерв (XII) — також руховий, він іннервує всі м'язи язика. Однобічне ураження цього нерва чи його ядра супроводжується порушенням функції язика. Крім ядер черепних нервів, особливістю структури довгастого мозку є наявність у ньому потужної маси нервових елементів, які не отримують аферентних волокон безпосередньо з периферії і не надсилають туди рухових волокон. Це сітчастий утвір (ретикулярна формація), який має нервові зв'язки тільки з іншими відді Будова і функції центральної нервової системи 307 лами ЦНС і є фактично однією з її иад-сегментарпих структур (див. с. 308). До иадсегмеитарних структур належать також ядра провідних шляхів ЦНС, що проходять через довгастий мозок. Основними з них є тонке і клиноподібне ядра (Голля і Бурдаха), а також оливні ядра. Функції довгастого мозку. У довгастому мозку розміщеш найважливіші автономні й соматичні центри, завдяки яким він виконує три основні функції — провідникову, рефлекторну і топічну. Провідникова функція довгастого мозку полягає в тому, що він з'єднує головний мозок зі спинним і проводить сигнали від одного відділу до іншого в обидва боки. Крізь нього проходять всі висхідні та низхідні шляхи спинного мозку, тут формуються присіпково-спинномозкові і сітчас-то-спиииомозкові шляхи і закінчуються кірковобульбарні волокна. Рефлекторна функція. Довгастий мозок бере участь у рефлекторній регуляції основних автономних (вегетативних) функцій, а також у ньому замикається низка сегментарних рефлексів. Автономні центри довгастого мозку і виконувані ними рефлекси поділяються па сегментарні (слиновидільний, смоктання, ковтання, чхання, кашлю тощо) і надсег-ментарні. Останні центри діють опосередковано, через інші структури ЦНС. Такими є дихальний і судиноруховий центри, які виконують життєво важливі функції, тому є постійно або ритмічно активними протягом усього життя організму. Дихальний центр. Якщо у тварини зробити перетин вище довгастого мозку, то вона може дихати, але якщо зруйнувати або охолодити довгастий мозок чи відокремити його від спинного мозку, дихання припиняється. Отже, ритмічні скорочення дихальних м'язів зумовлюються иейронними структурами довгастого мозку. Вважають, що ритм дихання формується в иейронних структурах бічної частини довгастого мозку, а структури нри-середиьої зони регулюють параметри дихання у разі зміни умов існування, забезпечуючи належний рівень легеневої вентиляції. Дослідження властивостей дихальних нейронів довгастого мозку свідчать про те, що вони здатні змінювати свою активність під впливом різних чинників: подразнення рецепторів, нервів і центральних структур, зміни концентрації 02 і С02, а також рН крові. У регуляції дихання бере участь і міст, де розміщений пневмотаксичний центр, який модулює функцію дихального центру, змінюючи частоту і глибину дихання. Докладніше про локалізацію, будову та принципи функціонування дихального центру див. с. 124. Судиноруховий центр також розміщений у довгастому мозку, що було встановлено Ф. В. Овсяпніковим у 1871 р. Вважали, що він знаходиться у ділянці дна IV шлуночка, і лише у 80-ті роки минулого століття вчені дійшли згоди, що головні структури судинорухового центру розміщені па протилежному боці—у иередиьобічиій ділянці довгастого мозку. Тут біля самої поверхні мозку розміщені скупчення нейронів (зони М і 5), електричне чи хімічне подразнення яких спричинює підвищення артеріального тиску, а руйнування — навпаки, максимальне зниження його. Трохи каудальніше розміщена зона Ь, нейрони якої чинять гальмівний вплив на зону 5. Судиноруховий центр тонічно активний — його нейрони перебувають у етапі постійного збудження, завдяки чому в організмі весь час підтримується нормальний рівень артеріального тиску. Ще одна важлива функція центру полягає у здійсненні судинорухових рефлексів. Барорецептори рефлексогенних зон кровоносної системи (дуги аорти, сонних пазух, серця тощо) надсилають інформацію до судинорухового центру про відхилення рівня артеріального тиску від нормального. З центру через спинний мозок і симпатичні нерви та через блукаючий нерв надходять імпульси, що змінюють судинний тонус і роботу серця, що сприяє відновленню нормального артеріального тиску. (Детальніше про функцію судинорухового центру див. с. 92.) Сегментарні рефлекси обмежуються невеликою кількістю пере- 308 ФІЗІОЛОГІЯ НЕРВОВОЇ СИСТЕМИ важно рухових ефекторів двох функцій: травної і захисної. До рефлексів травної системи належать слиновидільний, смоктальний, жувальний і ковтальний. їхні центри розміщені в ядрах ІХ-ХИ пар черепних нервів, а також лицевого і трійчастого нервів. Через ці центри здійснюються ланцюгові рефлекси, пов'язані з жуванням і ковтанням. Так, жування забезпечується координованою функцією жувальних м'язів, язика, щік, иідпебішія, дна порожнини рота, яку контролюють ядра трійчастого і під'язикового нервів. Коли харчова грудка, змочена слиною, дотикається піднебіння, основи язика чи слизової оболонки глотки, збудження рецепторів цих структур передається через чутливі волокна язико-глоткового нерва до центру ковтального рефлексу довгастого мозку, а звідти імпульси прямують через рухові волокна під'язикового, трійчастого, язикоглоткового і блукаючого нервів до м'язів порожнини рота, глотки, гортані та стравоходу. Внаслідок координованої діяльності цих м'язів здійснюється акт ковтання з одночасним перекриванням входів до носової частини глотки й гортані. Подібно до цього відбувається і смоктальний рефлекс. Центри згаданих рефлексів координують функцію жувальних м'язів щелеп, м'язів щік, язика, піднебіння, гортані, глотки і стравоходу між собою та з іншими центрами, наприклад з дихальним центром ири здійсненій смоктального чи ковтального рефлексів. Чхальний рефлекс і рефлекс кашлю є захисними відносно дихальної системи. Механічне чи хімічне подразнення рецепторів слизової оболонки носа через аферентні волокна трійчастого нерва збуджує чхальний центр, який анатомічно і функціонально перебуває у тісному зв'язку з дихальним центром. Звідти через відповідні нерви до м'язів рота, лиця, гортані та міжребрових м'язів надходять команди, що викликають координований чхальний рефлекс. Подібним чином відбувається рефлекс кашлю, але спричинюється він подразненням рецепторів дихальних шляхів — гортані, трахеї, бронхів. Блювотний рефлекс виконує захисну функцію відносно травної системи і всього організму. Виникає у випадках переповнення шлунка, потрапляння у кров шкідливих токсичних речовин. Із рецептивного поля рефлексу (корінь язика, глотка, слизова оболонка шлунка) сигнали надходять до блювотного центру аферентними волокнами блукаючого та язикоглоткового нервів. Шкідливі речовини з травного каналу кров'ю переносяться до головного мозку і безпосередньо подразнюють блювотний центр. Акт блювання відбувається внаслідок аптиперистальтичпих скорочень шлунка, а також скорочень різних м'язів черевної стійки і діафрагми. Тонічна функція довгастого мозку полягає у підтриманій певних груп нейронів чи відділів ЦНС у стані постійного збудження. Особливо яскраво ця функція виявлена щодо судинорухового центру і симпатичних нервів, які перебувають під його контролем. Довгастий мозок здійснює постійний тонічний вилив і на спинний мозок. Це підтверджує факт розвитку спінального шоку у тварин після відокремлення спинного мозку від довгастого. У всіх цих випадках топічна функція довгастого мозку найтісніше пов'язана з сітчастим утвором. 11.4.3. СІТЧАСТИЙ УТВІР Сітчастий утвір (ретикулярна формація, іогтаїїо геіїсиїагіз) — це особливе скупчення нейронів; починається у спинному мозку, проходить через увесь стовбур головного мозку, має зв'язки з проміжним мозком, а також з корою великого мозку (див. мал. 157). Нейрони сітчастого утвору мають по 1-6 довгих розгалужених дендритів і аксон, дві гілки якого утворюють: одна — еферентні сітчасто-спипномозкові шляхи, а інша — висхідні шляхи. До нейронів сітчастого утвору підходить велика кількість колатералей від усіх специфічних висхідних шляхів, які проходять через стовбур мозку, а також від низхідних шляхів, що йдуть від вищих відділів мозку, в тому чис Будова і функції центральної нервової системи 309
лі від кори великого мозку й мозочка. Отже, за своєю нейрошюю організацією і зв'язками сітчастий утвір поєднує впливи різних мозкових структур. Майже всі нейрони сітчастого утвору є полісенсорними, тобто можуть реагувати на подразнення кількох модальиостей, наприклад, на подразнення шкіри та дію світла, на больові й звукові подразнення. Ці нейрони мають також високу хімічну чутливість і тому легко пригнічуються деякими лікарськими речовинами, особливо барбітуратами, які повністю блокують їх. Ця обставина робить неможливим дослідження сітчастого утвору під наркозом. Функція сітчастого утвору зумовлена особливостями її нейронів. Однією з таких особливостей є здатність до підтримання стійкої тонічної активності. Практично всі нейрони безперервно генерують імпульси з частотою 5-10 за 1 с. Різні аферентні сигнали підсумовуються з цією фоновою активністю, викликаючи в одних клітинах її збільшення, а в інших, навпаки, гальмування. Тонічна активність сітчастого утвору. Постійна фонова активність нейронів сітчастого утвору пояснюється, з одного боку, їх високою хімічною чутливістю до виливу гуморальних чинників, які постійно деполяризують клітинну мембрану і збуджують нейрони. З іншого боку, це визначається особливостями аферентних зв'язків сітчастих нейронів, па яких конвертують колатералі від величезної кількості різних чутливих волокон, що зумовлює постійне подразнення сітчастих нейронів імпульсами чутливих систем організму. Внаслідок цього сітчасті нейрони перебувають у етапі постійної активності, що й зумовлює безперервний топічний вплив сітчастого утвору на розміщені вище і нижче структури мозку. Дихальний і судиноруховий центри розміщені в межах сітчастого утвору довгастого мозку. Нейрони цих центрів займають порівняно невелику ділянку, розміщену в його задньому відділі. Функція решти сітчастого утвору довгий час була невідомою. Висхідні впливи сітчастого утвору. У 1949 р. Г. Мегун і Дж. Моруцці встановили, що подразнення певної частини сітчастого утвору має стимулюваль-ний вплив на функцію кори великого мозку, подразнення інших її ділянок — гальмівний. Так, якщо тварина перебувала у стані сну, то після подразнення збуджувальної частини сітчастого утвору вона прокидалася і поводилась активно. Одночасно у мозку виникала реакція десинхронізації, яка охоплювала значні ділянки кори, що свідчить про генералізований характер висхідних активаційних виливів сітчастого утвору. Реакція десинхронізації не єдиний прояв впливу сітчастого утвору па функцію кори великого мозку. Відомо, що ири будь-якому аферентному збудженні у відповідних проекційних полях кори з'являється своєрідна електрична реакція, яка дістала назву первинної відповіді. 310 ФІЗІОЛОГІЯ НЕРВОВОЇ СИСТЕМИ Невдовзі за первинною відповіддю виникають додаткові коливання електричного потенціалу, які мають більший латентний період і генералізовапий характер. Це реакції вторинної відповіді. Спеціальні дослідження показали, що вторинні відповіді є електричним проявом надходження аферентного збудження до кори великого мозку за допомогою сітчасто-кіркових волокон. Отже, ири аферентному збудженні імпульсація через стовбур головного мозку і таламус потрапляє до кори великого мозку, що реєструється у вигляді первинної відповіді у проекційній зоні кори. Ця реакція має невеликий латентний період. Одночасно частина аферентної імиульсації по колатералях відгалужується до сітчастого утвору, активуючи його. Висхідними сітчасто-кірковими шляхами імпульсація надходить до кори, але вже у вигляді вторинної (затриманої) відповіді, яка виникає з довшим латентним періодом. Ця реакція охоплює не лише проекційну зону, а й сусідні ділянки кори, викликаючи в них етап підвищеної збудливості. Сумарну електричну відповідь на аферентне збудження називають викликаним потенціалом. Сітчастий утвір може виявляти й гальмівний вплив на ділянки ЦНС, розміщені вище, хоча організація гальмівної висхідної системи з'ясована не до кіпця. Низхідні впливи сітчастого утвору. Полегшувальні й гальмівні імпульси від сітчастого утвору проводяться також до спинного мозку сітчасто-сиииномоз-ковим шляхом (див. мал. 157), регулюючи його функцію. Так, подразнення сітчастого гігантоклітиииого ядра у довгастому мозку зумовлює пригнічення рефлекторної активності спинного мозку, причому це пригнічення є дуже глибоким і тривалим, навіть у тому випадку, якщо подразнення цього ядра спричинене лише кількома стимулами. Це дифузне неспецифічне гальмування охоплює майже всі рефлекторні дуги і ті нейрони, які беруть участь у передачі висхідної інформації, що призводить не лише до пригнічення рефлекторної діяльності спинного мозку, а й викликає часткову сенсорну депривацію кори великого мозку. У 1862 р. І. М. Сєчеиов виявив факт пригнічення спіпальних рефлексів ири подразненні стовбура головного мозку. Це було відкриттям центрального гальмування і одночасно сітчасто-спипномозкової системи. Проте механізм цього гальмівного ефекту вдалося розкрити лише майже через 100 років — у 50-ті роки XX ст. завдяки дослідженням Г. Мегупа, який показав, що локальне електричне подразнення гігаптоклі-тиниого ядра довгастого мозку викликає неспецифічне гальмування згинальних і розгинальних спинномозкових рефлексів. Механізм еєчеиівського гальмування виявився у тому, що збудження нейронів сітчастого утвору, яке поширюється через сіт-часто-сниппомозковий шлях, активує гальмівні вставні нейрони спинного мозку, аксони яких утворюють гальмівні синапси на а-мотонейропах. При цьому мембрана а-мотонейропів гіперноляризується, а їхня збудливість відповідно знижується. Крім гальмівного сітчастий утвір може виявляти й полегшувальний вплив на функцію спинномозкових структур. Центр полегшувальної системи сітчастого утвору знаходиться у ділянці моста і середнього мозку. Сітчастий утвір як прямо, так і опосередковано взаємодіє майже з усіма структурами і системами мозку, впливаючи па різноманітні функції. Одна з найголовніших — це функція неспецифічного підвищення збудливості, пов'язана з процесами активації та регуляції рівня притомності й уваги. Сітчастий утвір має також системи нейронів зі специфічними властивостями. Це сітчасті ядра, блакитне місце середнього мозку, які мають відношення до складних иоведіикових реакцій, павчашія, нам'яті, регуляції процесів активації та деяких вегетативних функцій. Сітчастий утвір тісно пов'язаний з гіпоталамусом і структурами лімбічної системи, які відіграють важливу роль в емоційно-мотиваційпих реакціях, а також з корою великого мозку. Будова і функції центральної нервової системи 311 1 1.4.4. ЗАДНІЙ МОЗОК Задній мозок у хребетних тварин представлений двома філогенетично і функціонально пов'язаними структурами — мостом і мозочком. У процесі еволюції мозочок відділився від решти мозку і перетворився па анатомічно відносно автономну структуру, пов'язану зі стовбуром мозку трьома парами ніжок. У зв'язку з цим будова і функції мозочка розглядаються, як правило, окремо. Міст (вароліїв) з'являється лише у ссавців, що пов'язано з розвитком кори півкуль великого мозку. Саме через міст проходять кірково-мозочкові шляхи. Міст розміщений між довгастим і середнім мозком, займає у людини передню частину заднього мозку і складається з двох структурио-фуикціоиальиих утворів: основної частини і покриву моста. В основній частині моста проходять кірково-сииппомозкові шляхи, між якими розміщуються власні нервові волокна моста, що прямують до мозочка. З основи моста виходять нерви, які зв'язують його певні зони з корою великого мозку. У ділянці покриву моста розміщені ядра сітчастого, ушору,. а також частково при-сіпково-завиткового нерва. Якщо нейрони ядер завитки сприймають сигнали від рецепторів спірального органа завиткової протоки, а нейрони верхнього присіпкового ядра — від рецепторів ирисіика і півколо-вих каналів, то присереднє і бічне присінкові ядра стосуються контролю рухових функцій, їхні нейрони дають початок ирисіи-ково-спииномозковому шляху, аксони якого досягають рухових нейронів спинного мозку. У ділянці покриву також розміщені рухові ядра трійчастого, лицевого та язико-глоткового нервів. Вони іннервують м'язи лиця, глотки. До цієї самої групи функціонально входять і рухові ядра черепних нервів довгастого мозку, які іннервують м'язи шиї. Вони беруть участь у здійсненні рухових рефлексів, спрямованих на відновлення і збереження постави. Рефлекси заднього мозку. У задньому мозку замикаються дуги бага
тьох вегетативних і соматичних рефлексів (мал. 158). Його функції найтісніше пов'язані з функцією присіпкового апарату (див. с. 376). До вегетативних належать рефлекси, що регулюють за допомогою парасимпатичних ядер лицевого і язикоглоткового нервів функцію слинних залоз. Крім того, ядра моста беруть участь у регуляції сльозо-і потовиділення, координації актів жування і ковтання, кліпання очима тощо. Парасимпатичне ядро блукаючого нерва входить до системи рефлекторної регуляції дихання і кровообігу. Соматичні рефлекси заднього мозку поділяють на ланцюгові й тонічні. Першу групу рефлексів розглянуто вище (див. с. 308). Нагадаємо про те, що в їх здійсненій беруть участь структури заднього мозку, зокрема нейрони ядер трійчастого і лицевого нервів. Тонічні, або постуральні (фр. розвите — положення тіла, поза), (статичні) рефлекси — це група рефлексів, у здійсненні яких беруть участь структури довгастого, заднього і середнього мозку. Вони спрямовані на підтримання певного положення тіла (пози) чи його частини у просторі. Залежно від джерела аферентних сигналів топічні рефлекси поділяють на шийні і присінкові. Ш и й н і топічні рефлекси викликаються збудженням ироиріорецеиторів м'я- 312 ФІЗІОЛОГІЯ НЕРВОВОЇ СИСТЕМИ зів шиї. Вперше вони були описані Р. Маг-ну сом у котів з перетином стовбура головного мозку. У чистому вигляді ці рефлекси можна спостерігати лише після руйнування присіпкового апарату, який надає відповідну інформацію про положення голови у просторі. Як показали досліди Р. Магпуса, закидання котові голови назад супроводжується зростанням тонусу м'язів — розгиначів передніх кінцівок з одночасним зниженням тонусу м'язів — розгиначів задніх кінцівок (мал. 159). Згинання голови викликає протилежні реакції, а повороти голови праворуч і ліворуч стосовно поздовжньої осі зумовлюють компенсаторне підвищення тонусу м'язів — розгиначів тих кінцівок, у бік яких повернено голову. Це рефлекси обертання за Р. Магпусом. Ефекторпою лапкою шийних рефлексів можуть бути і м'язи очного яблука, оскіль
ки вони рефлекторно забезпечують компенсаторне встановлення очей зі зміною положення голови. Усі шийні рефлекси є полісипаитич-иими. Імпульси від иронріорецепторів шийних м'язів проходять через клиноподібне ядро (Бурдаха) до відповідних центрів довгастого мозку, а звідти до рухових нейронів спинного мозку або рухових ядер відвідного нерва і викликають зміни положення тіла і тонусу м'язів очного яблука. Присінкові (вестибулярні) рефлекси поділяють па статичні і статокінетичпі. Статичні рефлекси пов'язані переважно зі збудженням рецепторів перетинчастого лабіринту нрисінка і забезпечують підтримання пози й рівноваги тіла при змінах його положення у просторі — рефлекси пози, рефлекси випрямлення. Наприклад, якщо кіт падає з висоти спиною донизу, то спочатку у нього завдяки присіпковому рефлексу випрямлення відновлюється нормальне положення голови (мордою донизу). Ця зміна положення голови збуджує пропріо-рецептори шийних м'язів, спрацьовує шийний рефлекс випрямлення, внаслідок чого нормального положення набуває також тулуб. Статокінетичпі рефлекси спрямовані на підтримання пози у разі зміни швидкості руху. Ці рефлекси зумовлені збудженням рецепторів півколових каналів, яке виникає під час руху ендолімфи всередині каналів. Прискорення під час обертання тіла у горизонтальній площині збуджує рецептори горизонтального півколового каналу і викликає рефлекторну реакцію окорухового апарату — горизонтальний очний ністагм. Ця реакція полягає в тому, що в момент прискорення обертання очні яблука рухаються в бік, протилежний напрямку обертання. Досягнувши максимально можливого відхилення, очні яблука швидко переміщуються у напрямку руху, і в поле зору потрапляє інша точка простору.
|