ЛЕКЦІЯ № 9 6 страница
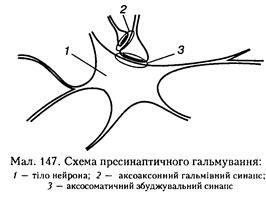
вових закінченнях (терміналях) збуджувальних нейронів (мал. 147). Медіатор, що виділяється в цих синаисах (припускається, що це ГАМК), викликає деполяризацію нервових закінчень за рахунок збільшення проникності мембрани для СГ. Іншою можливою причиною такої деполяризації може бути підвищення зовнішньої щодо нейрона концентрації К+ при тривалій активації аферентних входів. А гальмування збуджуваного нейрона виникає тому, що ця иреси-наптичиа деполяризація спричинює зменшення амплітуди ПД у иресииантичному закінченні, що, в свою чергу, призводить до зменшеного виділення квантів збуджувального медіатору (иейротрапсмітера), тому амплітуда ЗПСП знижується, і активація збуджуваного нейрона не відбувається. Пресинаптичне гальмування значно триваліше (сотні мілісекунд) за постсинаптичне. Воно особливо ефективне для обробки інформації, яка надходить до нейрона різіш-ми пресинантичиими шляхами, оскільки в цьому разі збудження, що надходить через один із входів, може бути вибірково зменшене або навіть повністю пригнічене, якщо немає інших виливів на інші сииантичиі входи. Припускають, що иресшішітачие гальмування пригнічує сигнали, біологічне значення яких у даний момент є незначним, і пропускає інформацію, що є найважливішою для організму. Песимальне (лат. реззітит — найгірший) гальмування. Про нього вже йшлося під час розгляду передачі збудження у нервово-м'язовому синаисі (див. с. 279). М. Є. Введепський вважав, що песимальне гальмування виникає внаслідок зниження функціональної лабільності збудливої структури в разі надмірних за силою або частих подразнень і виявляється пригніченням рефлекторної діяльності. Песимальне гальмування відіграє важливу роль у механізмах координації рефлекторної діяльності в ЦНС. Воно не пов'язане з активацією спеціалізованих гальмівних утворів, а виникає внаслідок поступового зниження збудливості, і ті самі імпульси, що раніше викликали рефлекторний акт, тепер його гальмують. Значення процесів гальмування у функціонуванні нервової системи надзвичайно Загальна фізіологія центральної нервової системи велике: воно обмежує пеконтрольоваие поширення збудження у ЦНС, забезпечуючи чіткі координовані реакції; відіграє охоронну роль, захищаючи нейрони від ушкоджешія внаслідок надмірних за силою подразнень. Значний внесок у вивчення механізмів центрального гальмування зробили праці таких учених, як І. М. Сєчеиов, Ч. Шерріпгтон, М. Є. Введеиський, Дж. Екклс, П. Г. Кос-тюк, П. М. Сєрков. Реципрокне гальмування, як один із видів складної координації рефлекторної діяльності, див. с. 296. 11.3.1.3. Координація рефлекторної діяльності Координація рефлекторної діяльності відбувається на рівні нервових центрів, що забезпечує узгоджену діяльність нервової системи в цілому. Розрізняють кілька основних принципів такої координації. Конвергенція (лат. сопуег§еге — зближуватися, сходитися) — це процес, коли збудження від кількох аферентних нейронів спрямовується па один ефектор-иий нейрон. Унаслідок цього допорого-ві ЗПСП можуть підсумовуватися па тілі цього нейрона, і локальний потенціал, що виникає, може досягати порогового рівня, необхідного для генерації ПД (мал. 148, а). Явище конвергенції лежить в основі просторової сумації збудження, завдяки чому полегшується виникнення відповідної рефлекторної реакції. Конвергенція може виявлятися також при взаємодії па одному нейроні збуджувальних і гальмівних сигналів від кількох різних джерел. Дивергенція (лат. (1іуег§епгла — розходження) збудження — здатність одного нейрона утворювати багато синаитичпих зв'язків з різними нейронами (див. мал. 148, б). Один і той самий нейрон може брати участь у різних реакціях і контролювати велику кількість нейронів, забезпечувати широкий перерозподіл імпульсів, що зумовлює іррадіацію збудження. Оклюзія (лат. оссіизіо — закривати, замикати). Суть оклюзії полягає в тому, що під час одночасного подразнення двох аферентних волокон, які частково конвертують на одних і тих самих нейронах, кількість збуджених нейронів виявляється меншою (на мал. 148, б їх 5), ніж сума збуджених нейронів при окремому подразненні кожного аферентного входу (4 + 4). В основі оклюзії лежать обидва процеси — конвергенції і дивергенції. Як показано на малюнку, розходження відростків кожного з двох аферентних нейронів па 4 еферентних поєднується з одночасним сходженням закінчень двох аферентних нейронів па кожному з 5 еферентних нейронів. Унаслідок взаємодії сигналів у такій системі відбувається оклюзія — часткове перекривання збудження, що виявляється зменшенням рефлекторної реакції у разі надмірного аферентного збудження.
296 ФІЗІОЛОГІЯ НЕРВОВОЇ СИСТЕМИ
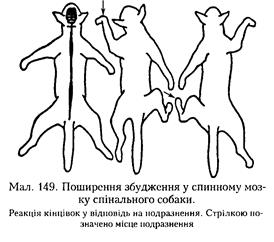
Спільний кінцевий шлях. Багато еферентних нейронів залучається до здійснення різних рефлекторних реакцій організму. Так, рухові нейрони, які іннервують дихальні м'язи, крім забезпечення акту вдиху беруть участь у таких рефлекторних реакціях, як кашель, чхання тощо. Звідси виходить, що ті самі рухові нейрони можуть залучатися до різних рефлекторних дуг. На рухових нейронах спинного мозку, що іннервують скелетні м'язи, закінчуються волокна різних низхідних шляхів. Тому один і той самий руховий нейрон, через який здійснюються різні рефлекторні реакції, вважають їхнім спільним кінцевим шляхом (див. мал. 148, а). Поширення (іррадіація) збудження. Ступінь поширення збудження залежить від сили подразнення і функціонального стану нервового центру (мал. 149). У реальних умовах, незважаючи па досить часту мож-ливість широкої іррадіації у ЦНС, збуджешія фактично поширюється лише в певних напрямках, викликаючи чіткі рефлекторні реакції, обмежені гальмуванням. Гальмування рефлекторних реакцій відіграє виключно важливу роль у процесах координації рефлекторної діяльності. Види та основні механізми гальмування ми щойно розглянули, а їх участь буде проілюстровано на наступних формах координації. Реципрокне (лат. гесіргосиз — взаємний), спряжене (поєднане) гальмування є важливим механізмом координаційної діяльності. Якщо подразнювати кінцівку спінального собаки електричним струмом, виникає звичайний згинальний рефлекс. Проте це досить складна рефлекторна реакція, оскільки одночасно зі скороченням м'язів — згиначів кінцівки відбувається розслаблення її м'язів-розгипачів. В основі такої взаємодії м'язів лежить координація центрів спинного мозку: збудження центру згиначів через вставні нейрони спричинює одночасне гальмування центру розгиначів (мал. 150). Таке гальмуваїшя називають реципрокним (Ч. Шерріштоп). Контрастні зміни стану нервових центрів також відіграють важливу роль у механізмах координації рефлекторної діяльності. Вони виникають після припинення збудження або гальмування. До таких явищ відносять послідовну і одночасну індукцію, яка може бути позитивною або негативною. Ч. Шерріштоп показав, що якщо подразнювати шкіру собаки, викликаючи слабкий чухальпий рефлекс, а потім па фоні цього рефлексу завдати електричного подразнення кінцівці, то чухальпий
Будова і функції центральної нервової системи рефлекс гальмується. Проте цей рефлекс різко підсилюється після припинення дії гальмівного подразника. Це приклад позитивної послідовної індукції. Гальмування діяльності під впливом сторонніх подразників є прикладом негативної одночасної індукції. Явище віддачі і ритмічні рефлекси. Явище віддачі полягає у швидкій зміні одного рефлексу іншим рефлексом протилежного значення. Так, після припинення подразнення, яке викликає сильний згинальний рефлекс, настає різке розгинання зігнутої кінцівки. Це пояснюється розгальмовуванням центру розгинального рефлексу після закінчення згинання кінцівки. Завдяки такому механізму виникають ритмічні рефлекси, коли один рефлекс зумовлює виникнення іншого (ланцюгові рефлекси). До ритмічних рефлексів належать чухальпий, локомоторний тощо. Принцип зворотного зв'язку. Будь-який руховий акт, спричинений аферентним подразненням, починається збудженням рецепторів м'язів, сухожилків і суглобових сумок, від яких нервові імпульси прямують до ЦНС. Крім того, якщо рух контролюється зором, до пропріоцептивиих приєднуються також зорові імпульси. Такі аферентні імпульси, які виникають в організмі від різних органів, називаються вторинними аферентними імпульсами на відміну від тих, які первинно викликали рефлекторі шй акт. Вторинна аферентація має першочергове значення у забезпеченні координації рефлекторної рухової діяльності, оскільки імпульси від виконавчих органів, які прямують до ЦНС, інформують її про характер виконуваних рухів. Це дає змогу коригувати рухову активність (П. К. Анохін). У разі ушкодження аферентного зворотного зв'язку рефлекторна рухова діяльність порушується. Так, у хворих з ураженнями иропріоцентивної чутливості рухи, особливо ходіння, втрачають свою плавність і точність, мають вигляд поштовхів. 11.4. БУДОВА І ФУНКЦІЇ ЦЕНТРАЛЬНОЇ НЕРВОВОЇ СИСТЕМИ 11.4.1. СПИННИЙ МОЗОК Будова спинного мозку. Спинний мозок — циліндричний тяж, розміщений у хребтовому каналі, складається з сірої і білої мозкової речовини. Сіра речовина спинного мозку па поперечному перетині має вигляд метелика або літери "Н". Розрізняють передні й задні роги сірої речовини спинного мозку, з'єднані широкою перетинкою сірої речовини — центральною проміжною речовиною, через яку проходить центральний канал. Крім передніх і задніх рогів у грудному відділі та верхніх поперекових сегментах спинного мозку виділяють бічні роги сірої речовини (мал. 151). Передні роги сірої речовини утворені скупченням тіл переважно рухових нейронів, які утворюють у поздовжньому напрямку передні стовпи різної товщини. У задніх рогах (стовпах) сірої речовини містяться нейрони, які передають сигнали від чутливих нейронів. До них у складі задніх корінців прямують центральні відростки нейронів чутливих спинномозкових вузлів. У бічних рогах (стовпах) містяться центр симпатичної нервової системи і другі нейрони снишюмозково-мозочкового шляху. Біла речовина складається виключно з аксонів нейронів спинного й головного мозку, вузлів спинномозкових нервів, утворюючи канатики спинного мозку: передній,-задній і бічний. Деякі з цих волокон не виходять за межі синішого мозку, інші, довші волокла з'єднують спинний мозок з головним. Групи волокон, які виконують певну функцію, утворюють нервові провідні шляхи. У людини від спинного мозку відходить 31 пара спинномозкових нервів, за якими його поділяють на відповідну кількість сегментів. Спинномозкові нерви з кожного сегмента утворюються з двох частин: переднього (вентрального) рухового і заднього (дорсального) чутливого корінців. На 298 ФІЗІОЛОГІЯ НЕРВОВОЇ СИСТЕМИ
задніх корінцях є потовщення — чутливий вузол спинномозкового нерва (спінальиий ганглій), де розміщені тіла чутливих (аферентних) нейронів, периферичний аксон яких передає інформацію від периферичних рецепторів, а центральний аксон прямує до спинного мозку. Через передні корінці виходять рухові (еферентні) волокна — аксони рухових нейронів спинного мозку (мал. 151). Така функціональна поляризація корінців (закон Беюія — МажандІ) добре виявляється після їх перерізання: передніх корінців — паралічем м'язів, задніх — втратою чутливості відповідних ділянок тіла. Властивості нервових клітин спинного мозку. Усі нейрони спинного мозку складають дві великі групи: рухові нейрони і вставні (проміжні, іитернейрони). Останніх приблизно в ЗО разів більше, ніж перших. Рухові нейрони (мотонейрони), розміщені у передніх рогах (стовпах) спинного мозку і поділяються, в свою чергу, на два основних типи: а- і у-мотонейрони, які іннервують різні за функцією волокна скелетних м'язів. Рухові а-нейрони (а-мотоиейропи) — це великі нервові клітини з довгими дендритами. На тілі й дендритах цих нейронів розміщені 10-20 тис. сипапсів, більша частина яких утворена закінченнями вставних нейронів. Аксони а-мотоиейроиів мають високу швидкість проведення збудження — 70-120 м/с. Деполяризація, яка виникає в а-мотоией-ропах при активації збуджувальних синапсів, викликає ритмічні розряди ПД. Частота імпульсації пропорційна ступеню деполяризації, проте звичайно вона не перевищує 10-20 імп/с, що зумовлено тривалою слідовою гіперполяризацією, яка розвивається після кожного ПД. Рухові у-нейрони (у-мотопейроии) розсіяні серед а-мотонейропів, мають значно менші розміри (не більш як 30-40 мкм). Вони не контактують безпосередньо з первинними аферентними волокнами, але мо-посииаптичио активуються волокнами низхідних шляхів. Аксони у-мотопейропів мають невеликий діаметр, що зумовлює їхню малу швидкість поширення збудження (10-40 м/с). Крім того, для них властива незначна слідова гіперноляризація, що дає змогу їм розряджатися з частотою 300-500 імп/с. Вставні (проміжні, іитернейрони) нейрони спинного мозку — це різнорідна група нервових клітин. Вони мають си-иаитичиі контакти тільки з іншими нейронами і становлять переважну більшість нейронів спинного мозку. Загальними властивостями вставних нейронів є їх розмір, ще менший, ніжу-мотонейропів; вони майже не мають слідової гіиерполяризації, а отже, генерують збудження з частотою понад 1000 імп/с. Навіть у відповідь па поодиноке подразнення вставні нейрони зазвичай відповідають ритмічним розрядом. До вставних нейронів належать гальмівні клітини Ренто, які, як правило, активуються аферентними волокнами від м'язових рецепторів. Складні взаємозв'язки між а- і у-мото-иейроиами і різними типами вставних нейронів лежать в основі трансформації ритму в нервових центрах спинного мозку. Провідні шляхи спинного мозку. У задніх, бічних і передніх канатиках спинного мозку проходять пучки нервових волокон. Це провідні шляхи центральної нервової системи. Будова і функції центральної нервової системи Провідні шляхи ЦНС поділяються на довгі й короткі. У більшості нейронів спинного мозку їхні аксони, що виходять із сірої речовини, закінчуються в межах спинного мозку. Ці нейрони та утворені їхніми аксонами нервові шляхи називають пропріо-спінальними. Роль цих шляхів полягає у зв'язуванні різних сегментів спинного мозку, тобто вони беруть участь у здійсненні між-сегментиих рефлексів. Так, сильне подразнення однієї кінцівки спіиальиої тварини викликає згинання не тільки цієї кінцівки і перехресний розгинальний рефлекс (див. мал. 149), а й рухи інших кінцівок. Тому в разі тривалої стимуляції виникають ритмічні згинальні й розгинальні рефлекси у трьох нестимульоваиих кінцівках — саме так, як це відбувається під час переміщення тварин. Довгі волокна у спинному мозку поділяють на аферентні (висхідні, центрипе-тальиі) і еферентні (низхідні, центрифугальні) шляхи. Аферентні (висхідні) шляхи (мал. 152, а). Найбільшими за кількістю волокон висхідними шляхами є топкий (Гол-ля) і клиноподібний (Бурдаха) пучки, які утворюють задні канатики спинного мозку (див. мал. 152, а). Ці пучки волокон, що є відростками нейронів чутливих спинномозкових вузлів, проводять збудження від пропріорецепторів м'язів, рецепторів сухожилків, дотикових (тактильних) рецепторів шкіри і частково рецепторів внутрішніх органів. Тонкий пучок (Голля) починається у крижовій частині спинного мозку, а волокна, які приєднуються до нього вище грудних сегментів, утворюють клиноподібний пучок. Кожне аферентне волокно, потрапляючи у задній канатик, поділяється Т-нодіб-но па висхідну й низхідну гілочки, причому низхідні гілочки незабаром закінчуються, а висхідні піднімаються до рівня довгастого мозку, де контактують зі скупченнями нейронів, які утворюють тонке й клиноподібне ядра. Волокна топкого пучка (розміщені в задніх канатиках нрисе-редньо) несуть до топкого ядра аферентні сигнали від нижньої частини тіла (переважно нижніх кінцівок). Клиноподібний пучок (Бурдаха) несе сигнали до клиноподібного ядра від рецепторів верхніх кінцівок і верхньої частини тулуба. Аксони нейронів, розміщених у тонкому і клиноподібному ядрах, па яких закінчуються волокна заднього канатика, перехрещуються між оливами у довгастому мозку і прямують до таламуса протилежного боку. Цей пучок, утворений волокна-
300 ФІЗІОЛОГІЯ НЕРВОВОЇ СИСТЕМИ ми нейронів другого порядку, дістав назву медіальної петлі. Нарешті, клітини ядер таламуса (нейрони третього порядку) своїми відростками досягають кори великого мозку. Волокна топкого і клиноподібного пучків мієліиізовані, вони проводять збудження зі швидкістю 60-100 м/с. Цією системою проводиться інформація, яка дає змогу дуже точно визначати локалізацію подразнень та їх зміну за простором і часом. Це найважливіший шлях специфічної соматосенсорної системи. Усі інші висхідні шляхи утворені аксонами нейронів, розміщених у сірій речовині спинного мозку, тобто нейронів другого порядку. Першою ланкою цих шляхів, як і в попередньому випадку, є нейрони спинномозкових вузлів, аксони яких уже на рівні спинного мозку закінчуються на нейронах другого порядку. Насамперед, це бічний спинномозково-таламічний шлях, який забезпечує температурну і больову чутливість, а також передній спинномозково-таламічний шлях, що проводить сигнали від дотикових (тактильних) рецепторів. Швидкість проведеш 1Я збудження цими шляхами становить 1 — ЗО м/с. Центральні відростки нейронів чутливого спинномозкового вузла досягають нейронів задніх рогів спинного мозку. Аксони цих нейронів або відразу переходять на протилежний бік, або, пройшовши кілька сегментів па своєму боці, перехрещуються і входять у білу речовину протилежного (коптралатерального) канатика, проходять без перерви весь спинний мозок, стовбур головного мозку і закінчуються в ядрах таламуса. Нейрони третього порядку несуть інформацію до кори великого мозку. Наступні два шляхи забезпечують передачу переважно проиріоцептивиої інформації до мозочка. Задній спинномозково-мозочковий шлях (Флексига). Центральні відростки нейронів чутливого спинномозкового вузла досягають нейронів проміжного стовпа сірої речовини (стовп Кларка). Шлях, не перехрещуючись, досягає мозочка, де у прадавній його структурі — корі черв'яка розміщені наступні нейрони цього шляху, аксони яких закінчуються у корі мозочка. Цим шляхом передаються сигнали від рецепторів м'язів і сухожилків зі швидкістю близько 140 м/с. Передній спинномозково-мозочковий шлях (Говерса). Центральні відростки нейронів чутливого спинномозкового вузла досягають до нейронів ироміжпо-присеред-иього ядра протилежного боку спинного мозку. їхні аксони через довгастий мозок, передні ніжки та черв'як мозочка прямують до кори мозочка, перехрещуючись вдруге. Цими волокнами передається інформація від сухожилкових, шкірних та пропріо-рецепторів зі швидкістю до 120 м/с, що має велике значення для підтримання тонусу м'язів, координації рухів, збереження рівноваги тіла. Еферентні (низхідні) шляхи (див. мал. 152, б) зв'язують відділи головного мозку з ефекториими нейронами спинного мозку. Вони проводять імпульси від центрів довільних рухів у корі, підкіркових структур та різних рефлекторних центрів, стовбура мозку. Кірково-спинномозкові (пірамідні) шляхи починаються у корі иередцеитраль-иої звивини головного мозку. Вій утворений аксонами пірамідних нейронів (гіганто-пірамідиих, Беца), вперше описаних професором Київського університету В. О. Бе-цом. Аксони цього шляху (у людини близько 1 м завдовжки) спочатку проходять повз таламус і основні ядра стовбура головного мозку, утворюючи внутрішню капсулу. На рівні довгастого мозку більшість волокон (75-90 %) перехрещуються і далі спускаються у складі бічних канатиків, утворюючи бічний кірково-спинномозковий (пірамідний) шлях (мал. 153). Друга, менша частина волокон йде іпси-латеральио (на тому самому боці) у передніх канатиках спинного мозку (передній кірково-спинномозковий (пірамідний) шлях), заходить до шийної і грудної частий спинного мозку, ие досягаючи поперекових сегментів. Аксони кірково-спиппомозкових шляхів закінчуються в певних сегментах спинного мозку, причому ті з них, які не перехрещу Будова і функції центральної нервової системи 301 валися на рівні стовбура мозку, перехрещуються у тих сегментах спинного мозку, де вони контактують зі вставними або руховими нейронами. Слід зауважити, що у людини тільки невелика частина (до 10 %) аксонів цих шляхів закінчується безпосередньо па рухових нейронах, частіше ці волокна спочатку підходять до вставних нейронів кожного сегмента. Основною функцією кірково-сиииномоз-кових шляхів є передача імпульсів із кори великого мозку для виконання довільних рухів. Надійність здійснення цієї функції підвищується завдяки наявності прямого й бічного шляхів, які багато в чому дублюють один одного. Кірково-спинномозковий шлях у процесі еволюції з'являється лише у ссавців і найбільш розвинений у людини. Крім кірково-спилпомозкових існують ще кілька низхідних шляхів, які утворюють екстрапірамідну систему волокон. До неї належать присінково-спинномозковий (вес-тибулосніпальний), червоноядерио-спииио-мозковий (руброспіпальпий), бічний і нри-середиій сітчасто-спишюмозкові (ретикуло-снінальні) шляхи (див. мал. 152, б). Червоноядерно-спинномозковий шлях (Мопакова) утворюється аксонами нейронів червоного ядра середнього мозку. Відразу після виходу з ядра ці волокна повністю переходять на протилежний бік. Частина з них прямує до мозочка і сітчастого утвору, а інша частина — до спинного мозку. У спинному мозку волокна розміщуються у бокових канатиках поряд із бічним кірково-сниииомозковим шляхом і закінчуються на вставних нейронах відповідних сегментів. Локальна електрична стимуляція волокон червопоядерно-спипномозко-вого шляху супроводжується збудженням а- і у-мотонейропів м'язів-згиначів (через вставні нейрони). Головне призначення цього шляху — управління м'язовим тонусом і здійснення рефлекторної (мимовільної) координації рухів. Присінково-спинномозковий шлях утворюється волокнами нейронів бічного при-сіпкового ядра (Дейтерса) довгастого мозку. Це один із найдавніших нервових шляхів, яким передаються імпульси від нрисінко-во-завиткового органа (вестибулярного
апарату) і мозочка до рухових нейронів спинного мозку. Він чинить збуджувальний вплив на а- і у-мотоиейропи м'язів-розги-иачів і гальмівний — па рухові нейрони м'язів згиначів. Цей шлях забезпечує підтримання тонусу скелетних м'язів, рівноваги тіла, координацію рухів. У разі ушкодження виникає порушення координації рухів та орієнтування у просторі. 302 ФІЗІОЛОГІЯ НЕРВОВОЇ СИСТЕМИ Присередній і бічний сітчасто-спинномозкові шляхи починаються від нейронів сітчастого утвору моста і довгастого мозку (див. с. 309), закінчуються в сірій речовині спинного мозку досить далеко від рухових нейронів. Волокна сітчасто-спин-иомозкового шляху, що йдуть від довгастого мозку, збуджують а- і у-мотопей-ропи м'язів-згиначів і гальмують мотоиейрони м'язів-розгииачів, тоді як волокна від моста мають протилежний вилив. Обидва шляхи передають тонізуючий вилив сітчастого утвору па спинний мозок. Після їх перерізання у тварини розвивається спі-пальпий шок. Спінальний шок. У хребетних тварин, особливо ссавців, вищі відділи ЦНС здійснюють контроль за діяльністю спинного мозку. Цей процес називають церебральним домінуванням, і внаслідок цього функції спинного мозку обмежуються, як правило, простими рефлекторними реакціями. Повний перетин спинного мозку у тварин або травма у людини спричинює негайний двобічний параліч (параплегія) усіх довільних м'язових рухів, керованих сегментами синішого мозку, розміщеними нижче перетину. Чутливість частий тіла, іиперво-ваних від цих сегментів, втрачається (арефлексія). Артеріальний тиск у людини у випадках спінального шоку впродовж перших днів може знижуватися до 40 мм рт. ст., проте через кілька днів зростає до 70-80 мм рт. ст. У людини відновлення вегетативних рефлексів настає через 1~3 міс, а рухових — через 6-12 міс. (за належного лікування і догляду за хворим), але вони стають менш досконалими і гірше координованими, ніж у нормі. Проте чутливість і довільний контроль за ділянками тіла, що ішіервуються сегментами спинного мозку, розміщеними нижче його розриву, не відновлюються. Період пригнічення всіх рефлексів після травми (перетину) спинного мозку називають спінальним шоком. У тварин такий шок можна викликати імітацією перетину — локальним охолодженням чи анестезією шийної частини спинного мозку. Хоча механізм виникнення спінального шоку чітко ие встановлений, проте вважають, що основною причиною цього явища є порушення функції основних низхідних (еферентних) провідних шляхів спинного мозку. Внаслідок цього блокуються полегшувальні впливи з боку вищих відділів ЦНС па а- і у-мотонейроии і деякі вставні нейрони. Крім того, відбувається розгальмування гальмівних вставних нейронів спинного мозку. Усе це призводить до тяжкого пригнічення рефлекторної активності (арефлексія). Точно пе відомо, як відбувається відновлення функцій спинномозкових нейронів і чому у людини цей процес триває кілька місяців. Існують припущення, що підвищення збудливості нейронів спинного мозку може бути пов'язане з впливом гуморальних чинників (наприклад, С02) на них або множинним проростанням (спру-тингом) у спинний мозок нових нервових волокон. Рефлекси спинного мозку. Для дослідження рефлексів, що здійснюються спинним мозком, використовують спінальну тварину, тобто тварину з відокремленим від головного спинним мозком. У такої тварини після зникнення явищ спінального шоку можна спостерігати велику кількість як соматичних, так і вісцеральних рефлексів. Соматичні рефлекси — це рухові рефлекси кінцівок, тулуба чи його частин. Згинальний рефлекс можна спостерігати на сиіиальиій жабі, злегка затиснувши пінцетом пальці задньої ланки. Лапка згинається і відсмикується від подразника. Для отримання розгинального рефлексу натискають па підошву попередньо зігнутої лапки. Вона розгинається і відштовхує подразник. Потиральний (чухальпий) рефлекс характерний для тварин з волосяним покривом шкіри. Це значно складніший за механізмом рефлекс; у його здійсненні беруть участь велика кількість сегментів спинного мозку і м'язів. У результаті задня лапка спіпальпої жаби прямує саме до подразнюваної ділянки шкіри і багаторазовими потираннями намагається усунути подразник. Рефлекси розтягання, або власні міотатичпі рефлекси м'язів — це рефлекси, які ініціюються подразненням пропріо Будова і функції центральної нервової системи 303 рецепторів — рецепторів скелетних м'язів або їхніх сухожилків. Однією з найпростіших рухових реакцій у людини є колінний (сухожилковий) рефлекс. Він виникає, коли сухожилок чотириголового м'яза піддають короткочасному розтяганню легким ударом нижче иа-коліика. Розтягання м'яза викликає короткочасне збудження розміщених усередині м'язової тканини рецепторів (внутрішньо-веретенних м'язових волокон). Це призводить до стрімкого скорочення чотириголового м'яза і до різкого, паче здригання, розгинання йоги у колінному суглобі. Подібні рефлекси можна отримати, вдаряючи по ахілловому сухожилку, ири цьому скорочується литковий м'яз. М'язи передніх кінцівок також реагують на розтягання їхніх сухожилків короткочасним скороченням. Латентний період цих рефлексів дуже короткий — якщо брати до уваги лише центральну частину рефлекторної дуги та найближчі до спинного мозку ділянки аферентного і еферентного нервів (до 3 см) — не перевищує 1,5 мс (С. Окс). Це свідчить про мінімальну кількість нейронів і синапсів у складі його рефлекторної дуги: всього два нейрони і один синапс між ними. Такі дуги називають моносинаптичними. Внаслідок своєї простої будови вони погано гальмуються і координуються головним мозком. Тому людина, що стоїть, може впасти навіть після незначного удару по п'ятковому (ахілловому) сухожилку. Якщо у спінальиої тварини розтягати, наприклад, литковий м'яз, то він скорочуватиметься тим сильніше, чим більше його розтягати. Цей рефлекс розтягання відрізняється від попередніх рефлексів за своєю тривалістю: в цьому випадку виникає тонічне скорочення, яке триває доти, доки м'яз розтягується, тоді як після подразнення сухожилка виникає швидке фазне скорочення. Ще одна особливість рефлексу розтягання полягає у тому, що в разі сильного розтягання м'яза рефлекс гальмується, і м'яз раптово розслаблюється. Попри всі відмінності обидві групи рефлексів здійснюються за єдиним механізмом. По-перше, їх викликають імпульси від нронріорецепторів м'язів (нервово-м'язових веретен), які розтягаються одночасно із сухожилками, по-друге, тривалість цих рефлексів зумовлена тривалістю подразнення. У випадку сухожилкового рефлексу в аферентних волокнах від м'язових веретен виникає короткочасний синхронний залп імпульсів, що спричинює швидке скорочеи-ня-здриганпя цього м'яза. При тривалому розтяганні м'яза в тих самих рецепторах та їхніх аферентних волокнах генерується низькочастотна імпульсація протягом усього часу розтягання, яка й зумовлює тривале скорочення м'яза. Вважають, що підтримання тривалого скорочення м'яза під час його розтягання забезпечується, крім того, приєднанням до рухового нейрона полісинап-тичних рефлекторних дуг, а також, імовірно, інших видів нервово-м'язових веретен.
|