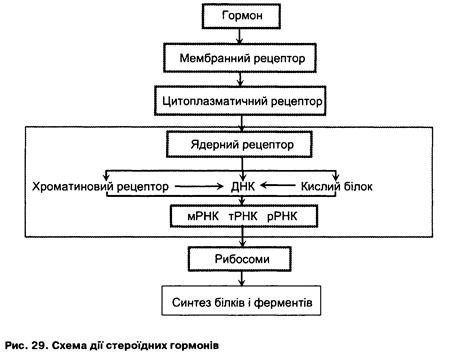
ЛЕКЦІЯ № 9 13 страница
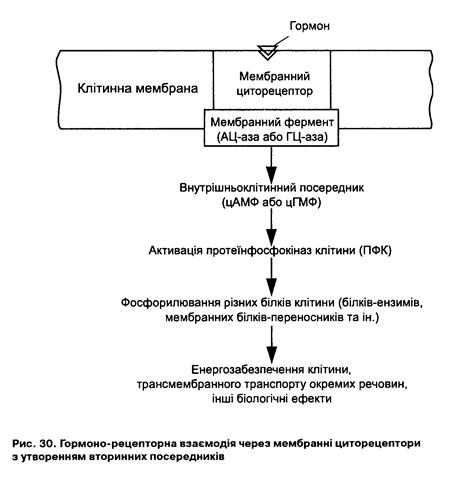
клітині (в тому числі ензимів) та зміна її функцій, відбувається диференціювання клітин. Пептидні гормони - це гідрофільні сполуки, які не здатні проникати через мембрану. Власне тому їх ефект зумовлений результатом взаємодії гормона з рецептором клітинної мембрани (рис. ЗО). Гормоно-рецепторна взаємодія запускає багато внутрішньоклітинних систем, причому деякі їх компоненти розташовані досить далеко від зовнішньої мембрани. Ця реакція починається з утворення біологічно активних речовин - вторинних посередників під дією комплексу "гормон-рецептор" специфічних ензимів. До вторинних посередників належать циклічні нуклеотиди (ц-АМФ, ц-ГМФ), кальцій та деякі продукти гідролізу фосфоліпідів (фосфоліпаза С-інозитол-три-фосфат - ІФз). Вони, змінюючи внутрішньоклітинний стан обміну речовин, швидкість і напрямок ферментативних процесів, а також проникність клітинних мембран, забезпечують специфічну дію гормону. Кількість мембранних рецепторів, так само як і внутрішньоклітинних посередників, непостійна, вона змінюється (модулюється) під впливом відповідних регуляторних механізмів.
Регулювання гормональної активності
Регулювання функцій синтезу і секреції гормонів ендокринними залозами здійснюється кількома шляхами:
1.Нейрогенне регулювання здійснюється у двох напрямках: а) прямий вплив нервової системи на синтез і секрецію гормонів (нейрогіпофіз, мозковий шар надниркових залоз); б) нервова система регулює гормональну активність опосередковано, змінюючи інтенсивність кровопостачання залози. 2.Гуморальне регулювання полягає у безпосередньому впливі на клітини залози концентрації субстрату, рівень якого регулює гормон (зворотний зв'язок). Так, рівень кальцію у крові впливає на секреторну активність прищитоподібної і щитоподібної залоз, де утворюються паратгормон і ти-рокальцитонін, а концентрація глюкози в крові визначає активність виходу інсуліну з підшлункової залози і т.д. 3.Нейрогуморальне регулювання здійснюється за допомогою гіпотала-мо-гіпофізарної системи. Функція щитоподібної та статевих залоз, кори надниркових залоз регулюється гормонами передньої частки гіпофіза (аденогіпофізом) - тронними гормонами: адренокортикотропним, тирео тропним, фолікулостимулювальним і лютеїнізивним. Дещо умовно до тропних гормонів відносять і соматотропний гормон гіпофіза, що здійснює свій вплив на ріст ще й опосередковано через гормон соматоме-дин, який утворюється у печінці. В аденогіпофізі також утворюється інтермедин (меланоцитостимулювальний гормон, МСГ) і пролактин, що здійснюють прямий вплив на периферійні органи. У свою чергу, вивільнення усіх зазначених гормонів аденогіпофіза залежить від гормональної активності нейронів медіальної ділянки гіпоталамуса. Тут утворюються гормони, що здійснюють стимулювальний чи гальмівний вплив на секрецію гормонів аденогіпофіза: рилізинг-фактори (ліберини) та інгібітори -статики. Ліберини і статини, впливаючи на продукцію тропних гормонів аденогіпофіза, регулюють активність деяких залоз внутрішньої секреції. Останнім часом використовують спільну назву - гіпофізіотропні гормони. На сьогодні встановлено шість гіпоталамічних рилізинг- та гальмівних гормонів: кортикотропін-рилізинг гормон (КРГ), тиротропін-рилізинг-гормон (ТРГ), гормон росту - рилізинг гормон (ГР-РГ), гормон росту інгібувальний гормон (ГРІ, або соматостатин), лютеїнізивний гормон -рилізинг гормон (ЛГРГ) або гонадотропін-рилізинг гормон (ГнРГ) і про-лактинінгібувальний гормон (ПІГ). Установлено, що екстракт гіпоталамуса проявляє пролактин-рилізинг-активність, отже, вважають, що існує і пролактин-рилізинг-гормон (ПРГ). Ділянка, в якій відбувається виділення гіпоталамічних рилізинг-гормонів та гальмівних гормонів, є серединним підвищенням гіпоталамуса. Вона містить небагато тіл нервових клітин, проте має чимало нервових закінчень, розміщених безпосередньо біля капілярних петель, з яких починаються портальні судини, що з'єднуються з гіпофізарною портальною системою судин. Функція значної кількості клітин, що синтезують гормони, регулюється за допомогою комбінації кількох названих механізмів. Крім того, багато гормонів, а також і гіпоталамо-гіпофізарні фактори впливають на утворення один одного. Тому, оцінюючи той чи інший механізм регуляції, необхідно мати на увазі, що мова йде лише про переважання того чи іншого з них. Регулювання гормональної активності більшості залоз внутрішньої секреції здійснюється за принципом від'ємного зворотного зв'язку: сам гормон (його кількість у крові) регулює своє утворення. Зазначений вплив опосередковується через утворення відповідних рилізинг-гормонів. Так, наприклад, за умови підвищення у крові рівня гормону кори надниркових залоз - кортизолу в гіпоталамусі вивільняється менше АКТГ-РГ, у результаті чого секреція гіпофізом АКТГ знижується. Це призводить до зменшення утворення кортикостероїдів наднирковими залозами і до зниження вмісту кортизолу в крові. Крім того, зворотний зв'язок у цій системі регулювання може бути опосередкований безпосередньо самими гормонами гіпоталамуса й аденогіпофіза. Названий механізм саморегулювання зберігається навіть за відсутності впливів на гіпоталамус з боку ЦНС (після повного відокремлення медіальної ділянки гіпоталамуса від інших відділів ЦНС). Але за нормальних умов інші відділи ЦНС беруть участь у пристосуванні цієї реакції до внутрішніх і зовнішніх потреб організму. Наприклад, утворення кортизо-лу корою надниркових залоз різко збільшується під час стрес-реакцій. Цьому передує збільшення секреції АКТГ-РГ і вихід АКТГ. Центральне регулювання гіпоталамо-гіпофізарної системи здійснюється центрами, що розташовані в передзоровому полі, у лімбічній системі та структурах стовбура мозку (довгастому і середньому мозку, мості). Сигнали від цих центрів до ядер гіпоталамуса передаються за допомогою нейронів, що належать до моноамінергічних систем головного мозку, трансмітерами в яких є біогенні аміни (норадреналін, дофамін, серотонін). До зазначених центрів ЦНС надходять не тільки нервові імпульси, але й інформація про рівень гормонів у крові завдяки наявності на мембрані нейронів зазначених відділів рецепторів, сприйнятливих до різних гормонів. Через моноамінові системи гормони впливають на специфічні структури гіпоталамуса і на продукцію ними РГ.
Методи дослідження ендокринних функцій
Експериментальні методи полягають у проведенні різних дослідів на тваринах. Серед них найчастіше застосовується метод видалення ендокринної залози (або її частини) і спостереження за зміною функцій організму - симптомами недостатності. Потім оцінюється замісна терапія шляхом уведення гормону в організм на тлі гіпофункції залози і спостереження за результатами цього введення. Можна одержувати результати і шляхом передозування - уведенням гормону на тлі наявної в організмі залози, яка нормально функціонує. Можна досліджувати порівняльний вміст гормону в крові, що надходить, і крові, що відтікає (як на рівні залози, так і будь-якого органа, де гормон продукується або використовується). У людини функція ендокринних залоз досліджується шляхом визначення концентрації гормонів у крові, а також швидкості виведення його з організму у складі біологічних рідин. Велику роль відіграють і дослідження пацієнтів із недостатньою функцією залози чи її гіперфункцією. Значне поширення одержав метод радіографії - уведення в організм радіоактивного ізотопу речовини, що входить до складу гормону (наприклад,13{]) і спостереження за розподілом цього ізотопу в тканинах організму.
9.2. Залози внутрішньої секреції
9.2.1. Гіпофіз Гіпофіз розташований у гіпофізарній ямці турецького сідла клиноподібної кістки. Через лійку з'єднується з гіпоталамусом. Його маса становить 0,5-0,7 г. Передню, проміжну і задню частки гіпофіза можна вважати сукупністю окремих ендокринних органів (у представників деяких тварин містять більше гормонально активних речовин). У людини проміжна частка -рудимент. Передня частка гіпофіза (аденогіпофіз) утворена епітеліальними поперечинами, між якими розташовуються синусоїдні капіляри. Серед клітин цієї частки виділяють більші - хромофільні аденоцити і дрібні -хромофобні аденоцити. Вузька проміжна частина утворена багатошаровим епітелієм, серед клітин якого виникають утвори, які нагадують пухирці, - псевдофолікули. Судинами лійки нейрогормони гіпоталамуса надходять у передню частку гіпофіза. Задня частка гіпофіза (нейрогіпофіз) утворена пітуїцитами - дрібними багатовідростковими клітинами і нервовими волокнами - аксонами клітин надзорового (супраоптичного) і пришлуночкових (паравентрику-лярних) ядер гіпоталамуса, що закінчуються на капілярах нейрогіпофіза. Цими аксонами (по надзорово-гіпофізарному і пришлуночково-гіпофізар-ному шляху) гормони, що продукуються у гіпоталамусі, транспортуються до задньої частки гіпофіза, а звідти надходять у кров.
Гормони передньої частки гіпофіза
Передня частка не пов'язана за допомогою нервових шляхів з ЦНС і тому її функціональна активність регулюється за допомогою нейрогормонів гіпоталамуса. Тут продукується дві групи гормонів: а) ефекторні гормони, що впливають на метаболічні процеси і регулюють ріст та розвиток організму; б) тропні гормони, що переважно регулюють секрецію інших ендокринних залоз. Гормон росту (ГР, соматотропін, СТГ) - прискорює ріст тіла, стимулює виділення інсуліноподібного фактора росту І (ІФР-І). Концентрація у крові непостійна: у дорослої людини коливається від 3 до 100 нг/мл сироватки крові. Викид його в кров збільшується під час глибокого сну, після м'язових вправ, за умови гіпоглікемії та інших станів організму. Ефект впливу ГР на хрящову тканину опосередковується через його дію на печінку, внаслідок чого в ній утворюються специфічні фактори (нині їх виділено не менше трьох), що називаються соматомединами, або поліпептидними факторами росту. Саме під впливом цих поліпептидних факторів відбувається стимуляція проліферативної і синтетичної активності хрящових клітин (особливо в зоні росту довгих трубчастих кісток, а також і в місці перелому кістки). Такий фактор, як соматомедин, виявляє мітотичну активність не тільки в хрящових, але і в інших клітинах. За своїм ефектом соматомедини близькі до інших стимуляторів проліферативної активності, таких, як фактор росту фібробластів, тромбо-цитарний фактор росту, ендотеліальний фактор росту, фактор росту нервів, тимозин, стимулятори кровотворення (утворюються у різних клітинах організму, докладніше про них - під час викладу відповідних розділів). Для кожного із зазначених факторів росту на мембранах відповідних клітин є рецептори, а їх вплив реалізується через посередників. Зміни в утворенні факторів росту зумовлюють порушення проліферації клітин. Наприклад, недостатнє утворення печінкових соматомединів може призвести до сповільнення зрощення кісток після переломів. Особливо велике значення ГР у період росту, коли концентрація його в крові підвищується. У цей період агоністами ГР є статеві гормони, збільшення секреції яких сприяє різкому прискоренню росту кісток (пубертатний скачок росту). Однак тривале утворення великої кількості статевих гормонів викликає протилежний ефект - припинення росту. Ріст дорослої людини може відновитися у разі надмірного зростання секреції ГР, що спостерігається, наприклад, на тлі пухлин гіпофіза. Тоді відновлюється проліферація клітин росткових зон, що призводить до акромегалії (гігантизму). За всіх зазначених вище станів вплив ГР супроводжується вторинною активацією обмінних процесів. Але гормон росту здійснює і прямий метаболічний вплив на жировий і вуглеводний обмін. Він бере участь у процесах ліполізу і підвищує стійкість клітин до гормону підшлункової залози інсуліну. Ефект антагонізму до інсуліну, що виражається у зниженні проникності клітин для глюкози, спостерігається за тривалої дії ГР, а одноразове введення його, навпаки, підсилює поглинання глюкози клітинами. Р'ЛІпотропін (Р-ЛПГ). Його фізіологічна роль точно не визначена. Вважають, що р-ЛПГ зумовлює розщеплення жиру з жирових депо і, як наслідок, збільшення рівня жирних кислот у крові. За структурою - це одношаровий поліпептид, що містить 91 залишок амінокислот у послідовності, яка характерна для структури ендорфінів та енкефалінів. Пролактин (ПЛГ) стимулює і підтримує утворення молока в грудних залозах, а також поведінку матері. До настання лактації грудна залоза формується під впливом жіночих статевих гормонів. Після пологів посилюється секреція гіпофізом пролактину і, як наслідок, виникає лактація - утворення та виділення молока грудними залозами. Крім того, пролактин зумовлює лютеотропну дію, тобто забезпечує функціонування жовтого тіла й утворення прогестерону в яєчниках. У чоловічому організмі він стимулює ріст і розвиток передміхурової залози. Адренокортикотропний гормон (АКТГ, кортикотропін) стимулює секрецію і ріст пучкової та сітчастої зон кори надниркових залоз. АКТГ зумовлює і пряму дію на тканини та органи. Він спричиняє розпад білка в організмі і гальмує його синтез (у цій якості він є антагоністом сомато-тропіну), знижує проникність стінки капілярів. Під його впливом зменшуються лімфатичні вузли, селезінка, щитоподібна залоза, знижується рівень лімфоцитів та еозинофілів крові. Тиреотропний гормон (ТТГ, тиротропін) стимулює виділення гормонів щитоподібною залозою, зумовлює збільшення її розмірів, кровона-повнення, розростання епітелію. ТТГ стимулює утворення білка тиреогло буліну в клітинах щитоподібної залози й активізує протеолітичні ферменти, під дією яких здійснюється його гідроліз і вивільнення гормонів щитоподібної залози. Гонадотропні гормони - фолікулостимулювальний (ФСГ) і лю-теїнізивний (ЛГ) є у чоловіків і в жінок. Вони стимулюють ріст і розвиток фолікулів у яєчниках і розростання сперматогенного епітелію у сім'яниках чоловіків. Ефекти ФСГ особливо значно виявляються в ембріональний період під час утворення фолікулів. В онтогенезі чутливість яєчників до ФСГ і Л Г підвищується у період статевої зрілості, коли обидва гормони забезпечують овуляцію та лютеїнізацію оваріальних фолікулів у жінок і виділення тестостерону - в чоловіків. Кортикотропоцити передньої та проміжної часток синтезують великий білок-попередник, унаслідок розщеплення якого виникає група гормонів. Після видалення сигнального пептиду утворюється прогормон - проопіомеланокортин (ПОМК). Синтезування ПОМК відбувається також у гіпоталамусі та інших частинах нервової системи, легенях, травному тракті і плаценті. У кортикотропоцитах цей гормон гідролізує до АКТГ, р-ЛПГ та невеликої кількості р-ен-дорфіну. У проміжній частці ПОМК далі гідролізує до кортикотропіно-подібного пептиду проміжної частки (КППП), у-ЛПГ і значної кількості Р-ендорфіну. Утворюються також а- і р-МТГ (а-, Р-, у- меланотропні гормони; загальна назва - меланотропін чи інтермедин). Поскільки проміжна частка - рудимент, то в дорослих а- чи р-МТГ інколи не виділяються. Меланоцитостимулювальний гормон (МСГ) підвищує секрецію меланіну в клітинах шкіри, спричиняючи її потемніння. Цей ефект добре виражений у земноводних. У дорослої людини МСГ у гіпофізі виявлений у дуже незначній кількості. Роль МСГ як гормону, що регулює забарвлення шкіри, у здорової людини ще не до кінця вияснена, хоча за деяких патологічних станів вона проявляється, зумовлюючи потемніння шкіри чи її окремих ділянок. Це спостерігається за первинної недостатності кори надниркових залоз (хвороба Адісона), його вторинної гіперактивності (хвороба Кушинга). Під час вагітності також може відбуватися потемніння окремих ділянок шкірних покривів (навколо сосків, слизових оболонок). Це зумовлено не тільки гіперсекрецією МСГ, але й АКТГ та інших гормонів гіпофіза, надниркових залоз і плаценти. Усіх їх поєднує те, що вони є похідними ПОМК і подібні один до одного за хімічною структурою. МСГ виявлено і в низці інших відділів мозку людини. Висловлено припущення про можливу роль його і близьких до нього сполук у регуляції росту та розвитку плода, у процесах, пов'язаних з пам'яттю.
Гормони задньої частки гіпофіза
Синтез гормонів нейрогіпофіза відбувається у надзоровому і пришлу-ночкових ядрах передньої ділянки гіпоталамуса. Потім вони у вигляді гранул транспортуються нервовими відростками, що контактують з кровонос- ними капілярами, які розташовані в задній частці гіпофіза. Тут гранули накопичуються, а потім з кров'ю розносяться до периферійних органів-міше-ней. Вазопресин (антидіуретичний гормон, АДГ) сприяє затримуванню рідини в організмі. У фізіологічних концентраціях цей гормон звичайно регулює вміст води і виділення її нирками, тобто він є одним з активних регуляторів осмолярності рідких середовищ організму, об'єму крові і рівня артеріального тиску. Якщо концентрація гормону в крові досить висока, то виявляється ще і його судинозвужувальний ефект. Сигналом до викиду гормону в кров є зниження артеріального тиску чи об'єму крові, що циркулює. Цей гормон також бере участь у реалізації механізмів сприйняття болю, антистресорних реакціях організму, тому названі впливи активують його викид у кров. Вазопресин впливає на клітини трьох типів: а) клітини ниркових канальців; б) непосмугованом'язові клітини кровоносних судин; в) клітини печінки. Діючи на нирки, він збільшує реабсорбцію води в їх ка-нальцях і цим зменшує сечоутворення. У кровоносних судинах гормон стимулює скорочення непосмугованих м'язових клітин і звужує судини, підвищуючи артеріальний тиск. У печінці стимулює гліконеогенез і гліко-геноліз. Окситоцин сприяє скороченню матки вагітних, зумовлює виділення молока, може також брати участь у процесі лютеолізу. Останній здійснюється шляхом впливу на високоспеціалізовані міоепітеліальні клітини, які вистилають протоки грудної залози. Під час лактації скорочення міоепітеліальних клітин зумовлює витискання молока з альвеол залози в молочний синус і виділення його через сосок. Виділення молока регулюється власне окситоцином. Крім названих гормонів в одній чи кількох частках гіпофіза виявлені інші поліпептиди: холецистокінін (ХЦК), гастрин, ренін, ангіотензин II, пептид, споріднений з геном кальцитоніну (ПСГК).
9.2.2. Надниркові залози Дві надниркові залози (права і ліва) розташовані на верхніх полюсах нирок. За формою вони нагадують сплющену піраміду зі злегка заокругленою вершиною. Надниркова залоза (масою 5-8 г кожна) складається з двох ендокринних утворів (один навколо іншого), представлених кірковою і мозковою речовиною. У корі надниркової залози розрізняють три зони змінної чіткості: клу-бочкову (зовнішню), пучкову (середню) і сітчасту (на межі з мозковою речовиною). Клубочкова зона утворена дрібними скрученими клітинами, що розташовані у вигляді клубочків, які переходять у колони клітин, що формують пучкову зону. Пучкова зона сформована великими світлими клітинами, що розташовуються довгими тяжами і заповнені краплями ліпідів. Внутрішня частина клубочкової зони переходить у сітчасту зону, де клітинні колони переплітаються у сітку. У сітчастій зоні дрібні клітини ут ворюють скупчення невеликих розмірів. Клубочкова зона становить 15% від маси надниркової залози, пучкова - 50%, сітчаста - 7%. Мозкова речовина (мозковий шар) утворена скупченнями великих круглих чи багатокутних клітин, які розділені синусоїдними капілярами. Ці клітини добре зафарбовуються солями хрому в коричневий колір і тому названі хромафінними. Насправді вона є симпатичним ганглієм, у якому постгангліонарні нейрони втратили аксони і стали секреторними клітинами. Клітини синтезують секрет під час збудження прегангліонарних нервових волокон, які досягають залози через вісцеральні нерви.
Гормони кори надниркових залоз
Кора надниркових залоз виділяє три групи гормонів: пучкова зона виділяє глюкокортикоїди (гідрокортизон, кортизон і кортикостерон) -стероїди з різноманітним впливом на метаболізм вуглеводів та білків; клубочкова - мінералокортикоїди (альдостерон, дезоксикортикостерон), необхідні для підтримання балансу натрію та об'єму позаклітинної рідини; сітчаста - статеві гормони (андрогени, естрогени, прогестерон) і, частково, глюкокортикоїди. Глюкокортикоїди свою назву отримали через здатність підвищувати рівень цукру в крові шляхом стимуляції утворення глюкози в печінці. Цей ефект є наслідком глюконеогенезу - дезамінування амінокислот у разі посилення розпаду білків. За такої умови вміст глікогену в печінці може навіть підвищуватися. Крім того, посилюється мобілізація жиру з депо і використання його для утворення АТФ. Кортизон впливає і на інші види метаболізму, що багато в чому визначається його рівнем у крові. Так, він може впливати навіть на мінеральний обмін, хоча для цього концентрація кортизону повинна бути набагато вищою, ніж основного мінералокортикоїду альдостерону. І взагалі, чим вища концентрація кортизону в крові, тим різноманітніший його вплив. Наприклад, у невеликій концентрації глюкокортикоїди активують, а у великій, навпаки, пригнічують імунні механізми організму. Високий рівень кортизону в крові зумовлює використання амінокислот для утворення глюкози і виявляє антианаболічну дію. Особливо значно знижується синтез білків м'язів, у такому разі може виникати і катаболічний ефект - розщеплення м'язових білків для вивільнення із них амінокислот. Глюкокортикоїди й АКТГ також впливають на нервову систему (збуджують її, спричиняють безсоння, ейфорію), на імунні та інші системи організму. Про комплексний вплив кортизону на різноманітні функції організму можна судити за такими змінами, що зумовлені його недостатністю: 1) гіперчутливістю до інсуліну; 2) зниженням запасів глікогену в тканинах; 3) зниженням активності глюконеогенезу; 4) недостатньою мобілізацією білків периферійних тканин; 5) ослабленням реакції жирових клітин на звичайні ліполітичні стимули; 6) гіпотензією; 7) затримкою росту; 8) м'язовою слабкістю і швидкою стомлюваністю; 9) зниженням здатності до посиленого виділення води в разі водного навантаження, 10) психічними та емоційними зрушеннями. У фізіологічних умовах зазначені метаболічні ефекти глюкокортикоїдів збалансовані. За необхідності вони швидко забезпечують потреби організму в енергетичному матеріалі. Тому під час гострих стресових станів гіпоталамо-гіпофізарно-надниркова система активується в першу чергу і рівень глюкокортикоїдов у крові підвищується. Під час повторної і тривалої дії стресового чинника реакція поступово згасає. Важливою властивістю глюкокортикоїдів є їх протизапальна дія, пов'язана з тим, що вони знижують проникність судинної стінки і блокують секрецію серотоніну, гістаміну, кінінів та систему плазмін-фібри-нолізин, а також пригнічують утворення антитіл. Протизапальна дія глюкокортикоїдів застосовується у клінічній практиці, наприклад, для лікування хворих на ревматизм. Розроблено низку мазевих препаратів для зовнішнього застосування (фторокорт, преднізолон тощо) У регуляції секреції глюкокортикоїдів бере участь гіпоталамус. У ядрах переднього гіпоталамуса продукується кортиколіберин, який через ворітну систему надходить до аденогіпофіза і сприяє синтезу АКТГ, що стимулює утворення кортикостероїдів. У свою чергу, продукування АКТГ залежить від рівня глюкокортикоїдів у крові (за механізмом негативного зворотного зв'язку) і від рівня гіпоталамічного рилізинг-гормону. Мінералокортикоїди беруть участь у регуляції мінерального обміну і водного балансу організму. Найактивнішим з них є альдостерон. Під його впливом посилюється реабсорбція натрію і зменшується реабсорбція калію у ниркових канальцях, а це призводить до затримки іонів натрію і хлору в організмі і до збільшення виділення іонів калію та водню. На відміну від глюкокортикоїдів, мінералокортикоїди посилюють розвиток запальних процесів за рахунок підвищення проникності капілярів і серозних оболонок. Вони також беруть участь у регуляції тонусу кровоносних судин і сприяють підвищенню артеріального тиску. Посилення синтезу і секреції мінералокортикоїдів здійснюється, по-перше, під впливом ангіотензину II, по-друге - під впливом АКТГ, що, у свою чергу, відбувається під впливом кортиколіберину гіпоталамуса. Інгібіторами синтезу і секреції мінералокортикоїдів є дофамін, передсердний натрій-уретичний гормон, значне підвищення концентрації іонів натрію у крові. Статеві гормони кори надниркових залоз мають значення для розвитку статевих органів у ранньому дитячому віці і для появи вторинних статевих ознак у той період, коли секреторна функція їх ще незначна. Крім того, естрогени проявляють антисклеротичний ефект (насамперед у жінок). Статеві гормони (особливо андрогени) сприяють обміну білків, стимулюючи їх синтез в організмі. Гормони мозкової речовини надниркових залоз
Мозкова речовина надниркових залоз виробляє катехоламіни: адреналін, норадреналін і дофамін, що виявляють метаболічні ефекти, які забезпечують постачання організму енергією. Оскільки основним джерелом енергії в організмі є глюкоза, катехоламіни індукують розщеплення глікогену в печінці та м'язах. Крім того, вони стимулюють ліполіз у жировій тканині і протеоліз у печінці, що теж сприяє енергетичному забезпеченню організму. Особливо важливі зазначені метаболічні процеси під час розвитку стресових ситуацій. Під впливом стресового чинника різко підвищується секреція катехоламінів, що спричиняє посилення серцевої діяльності, звуження судин внутрішніх органів і розширення судин, що постачають м'язи кров'ю, пригнічують перистальтику травного тракту, зумовлюють розширення бронхів. Усі ці зміни створюють оптимальні умови для боротьби організму із стресовими чинниками. Пептиди мозкової речовини надниркових залоз. Крім катехоламінів у мозковій речовині надниркових залоз синтезуються і пептидні гормони. Тут виявлені речовина Р, інтерстиціальний вазоактивний поліпептид, со-матостатин, (5-енкефалін. Функціональна характеристика цих сполук наводиться у відповідних розділах. Регулювання утворення гормонів мозкової речовини надниркових залоз здійснюється нервовою системою. Вона іннервуєтся прегангліонарними волокнами симпатичної нервової системи. Постгангліонарні нейрони відсутні, тому адреналін ще називають "рідким нейроном". Крім того, секреція катехоламінів контролюється задньою групою ядер гіпоталамуса.
9.2.3. Щитоподібна залоза Щитоподібна залоза масою 20-30 г розташована на шиї перед гортанню й охоплює її спереду і з боків. У ній розрізняють дві частки (праву та ліву) і перешийок. Попереду гортані від перешийка відходить пірамідна частка. Паренхіма залози складається з багатьох ацинусів - сферичних фолікулів, стінка яких утворена одним шаром тироцитів, що лежать на базальній мембрані. Кожен фолікул обплетений густою мережею кровоносних і лімфатичних капілярів. У порожнині фолікула міститься густа речовина білкової природи, забарвлена в рожевий колір, - колоїд щитоподібної залози (тиреоглобулін), що містить гормони. У стінках фолікулів між тироцитами і базальною мембраною, а також між фолікулами розташовані більші світлі парафолікулярні клітини (С-клітини).
Гормони щитоподібної залози
У фолікулах щитоподібної залози з тирозину утворюються два гормони: трийодотиронін (Тз) і тироксин (Т4). Трийодотиронін може також ут ворюватися у периферійних тканинах шляхом дейодування Т4. У пара-фолікулярних клітинах синтезується гормон кальцитонін. Регуляція утворення, як і гормональна активність, зазначених двох типів гормонів принципово відрізняється. Йодовані гормони. Біологічно активним гормоном є трийодотиронін, що утворюється переважно на периферії шляхом дейодування тироксину (в гіпофізі, печінці та нирках). Тироксин практично не володіє активністю. Надійшовши у кров, обидва йодовані гормони зв'язуються з одним із білків-переносників плазми крові й у такому вигляді транспортуються до органа-мішені. Утворення Т3 і Т4 регулюється ТТГ гіпофіза. У свою чергу, секреція ТТГ регулюється ТРГ гіпоталамуса. За механізмом негативного зворотного зв'язку тиреоїдні гормони впливають на гіпоталамус і гіпофіз; коли їх концентрація в крові стає максимальною, секреція ТТГ знижується до мінімуму. І навпаки, низька концентрація тиреоїдних гормонів у крові зумовлює високу швидкість секреції ТТГ. Рівень тиреоїдних гормонів у крові досить постійний, хоча секреція ТТГ трохи посилюється безпосередньо перед сном, а протягом ночі поступово зменшується. Характерні також коливання гормональної активності у зв'язку із сезонними змінами температури. Функції тиреоїдних гормонів. Тиреоїдні гормони беруть участь у диференціюванні клітин, що розвиваються, а в уже диференційованих клітинах регулюють обмінні процеси. Особливо важлива для організму участь тиреоїдних гормонів у процесах транскрипції і трансляції. Роль гормонів проявляється тим значніше, чим менший вік людини; краще це видно в разі гормональної недостатності. Так, в ембріональний чи ранній постна-тальний період гіпофункція щитоподібної залози виявляється у недорозвиненні мозку {кретинізм) і малому рості (карликовість). У дорослому організмі порушення продукції тироксину особливо помітно за умови зміни активності обмінних процесів, процесів теплоутворення: у разі гіпофункції хворий страждає від холоду, гіперфункції - від тепла.
|