Процесс трансдукции при зрении
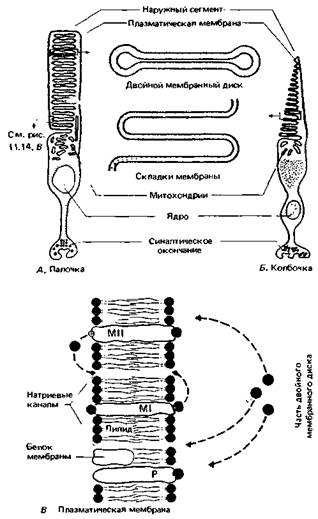
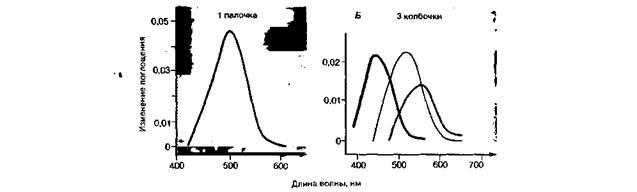
Структура фоторецепторов. У человека слой рецепторов сетчатки состоит примерно из 120 млн. палочек и 6 млн. колбочек (рис. 11.13). Плотность колбочек (их число на единицу площади) макси- 248 ЧАСТЬ III. ОБЩАЯ И СПЕЦИАЛЬНАЯ СЕНСОРНАЯ ФИЗИОЛОГИЯ мальна в середине центральной ямки, а палочек - вокруг этой ямки; в ней самой они полностью отсутствуют. Оба типа рецепторов сходны по структуре в том смысле, что и у тех, и у других есть наружный сегмент, состоящий примерно из тысячи мембранных дисков (палочки) или мембранных складок (колбочки). Он соединяется с остальной частью клетки узкой «ресничкой» (рис. 11.14, А, Б). На поперечном срезе сетчатки наружные сегменты фоторецепторов образуют правильную мозаику. В середине центральной ямки диаметр наружного сегмента колбочки составляет примерно 2 мкм, что соответствует углу зрения около 0.4'. К периферии сетчатки этот диаметр увеличивается. Зрительные пигменты. Молекулы зрительных пигментов очень упорядоченно включены в двойной липидный слой мембранных дисков наружных сегментов (рис. 11.14 ). У приготовленного в темноте раствора зрительного пигмента палочек (родопсина, или «зрительного пурпура») красный цвет, так как он особенно сильно поглощает зеленые и синие лучи. Более точно можно оценить эту способность, сняв спектр поглощения зрительного пигмента. У родопсина два его максимума - один в видимой части спектра (около 500 нм), другой в ультрафиолетовой (около 350 нм). Кривые поглощения зрительных пигментов одиночных фоторецепторов получают с помощью микроспектрофотометрни:иссекают кусочек сетчатки и под микроскопом направляют очень узкий пучок света с разной длиной волны сквозь изолированный наружный сегмент рецептора на высокочувствительный фотоэлемент (рис. 11.15). Этот способ показал, что: 1) у зрительных пигментов палочек и колбочек разные спектры поглощения; 2) спектр поглощения палочек такой же, как у родопсина, и достаточно близок к кривой спектральной чувствительности скотопического зрения (рис. 11.4). Родопсин состоит из гликопротеина (опсина) и хромофорной группы - 11- цис -ретиналя, т.е. альдегида витамина А (ретинола); 3) есть три типа колбочек,различающихся своими зрительными пигментами (рис. 11.15). Выцветание и регенерация зрительных пигментов в результате поглощения света. Процесс трансдукции в фоторецепторе начинается с поглощения фотона π-электронами сопряженных двойных связей ретиналя. В результате молекула переходит на более высокий энергетический уровень и испытывает более сильные колебания. При этом с вероятностью 0,5-0,65 («квантовая эффективность») происходит стереоизомеризация ретиналя. т.е. его переход из 11 -цис в полностью транс -форму. Затем вся молекула пигмента в несколько этапов разрывается с образованием в конечном счете ретинола и опсина (см. учебники биохимии или 1-е издание этой книги). Чтобы использовать данный первичный фотохимический процесс для передачи сигналов, он должен быть «превращен» в изменение мемб-
ранного потенциала клетки. Детали превращения еще не вполне ясны. Согласно одной из гипотез, при нем «активируются» связанные в мембранных дисках ионы кальция, которые в результате диффундируют из них в плазматическую мембрану наружного сегмента палочки или же в случае колбочек - из одного участка плазматической мембраны в другой (рис. 11.14, В). В итоге проводимость мембраны для мелких ионов, особенно натрия, уменьшается и возникает вторичный рецепторный потенциал, обсуждаемый ниже. Более поздняя гипотеза основана на представлении о сравнительно высокой натриевой проводимости мембраны наружного сегмента в темноте и, следовательно, прохождении через нее «темнового тока». Эта проводимость зависит от взаимодействия циклического 3'-5'-гуанидинмонофосфата (цГМФ) с белковыми молекулами натрие- ГЛАВА 11. ЗРЕНИЕ 249
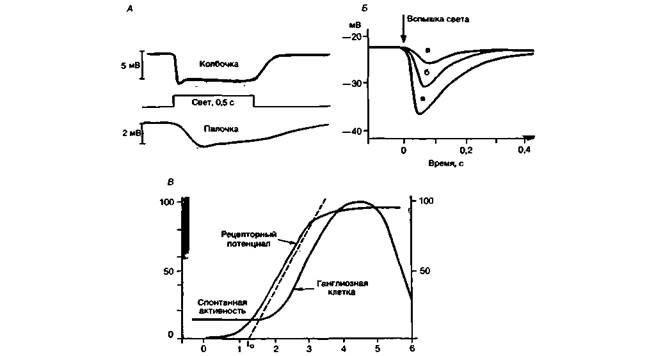
вых каналов мембраны рецептора; цГМФ заставляет их оставаться «открытыми». Тогда первичный фотохимический процесс состоит в восстановлении за несколько миллисекунд цГМФ за счет цепочки ферментативных реакций: натриевая проводимость уменьшается и возникает гиперполяризационный вторичный рецепторный потенциал. Предполагается, что при этом молекула родопсина, превратившись в метародопсин II за счет поглощения фотона, в крайне быстрой последовательности активирует множество молекул фермента («трансдуцина»), что приводит к начальному «усилению» первичного фотохимического процесса, поскольку «трансдуцин» управляет цГМФ [4, 5, 8, 34, 46, 53, 56а]. Первичный рецепторный потенциал фоторецепторов и фотохимические компоненты световой и темновой адаптации. Конформационное изменение молекулы родопсина генерирует с очень коротким латентным периодом (меньше 1 мс) электрический потенциал, называемый первичным (ранним) рецепторным потенциалом (РРП) (рис. 11.16). В нем несколько компонентов, которые можно выявить, постепенно снижая температуру сетчатки. Когда зрительный пигмент распадается на ретинол и опсин, происходит его ресинтез за счет цепи поглощающих энергию ферментативных реакций. Если на сетчатку падает постоянный поток фотонов, устанавливается динамическое равновесие между процессами выцветания зрительного пигмента под действием света и его регенерации с участием ферментов. Если сетчатки достигает меньше света, оно сдвигается в сторону более высокой концентрации родопсина. Это фотохимическая основа хорошо всем известной темновой адаптации глаза (см. с. 254, 255). В ходе нее амплитуда РРП, вызываемого световыми стимулами постоянной интенсивности, повышается, поскольку возрастает вероятность соударения падающих фотонов с молекулами невыцветшего родопсина [8]. Вторичный рецепторный потенциал. Если РРП генерируется за счет синхронных конформационных изменений молекул зрительного пигмента, то вторичный (поздний) рецепторный потенциал ПРП, не связанный с РРП непосредственно, - это изменение мембранного потенциала фоторецептора. В темноте он составляет от —25 до —40 мВ; под действием света происходит гиперполяризация. Амплитуда
этого рецепторного потенциала растет с увеличением интенсивности светового стимула (рис. 11.17), причем у палочек медленнее, чем у колбочек. Другими словами, временная инерция палочковой системы больше, чем у колбочковой (с. 256). Неодинаковая спектральная чувствительность вторичных рецепторных потенциалов различных типов колбочек (рис. 11.18) подтверждает результаты микроспектрофотометрии и говорит в пользу трихроматнческой (трехкомпонентной) теории цветового зрения (с. 271). т.е. присутствия трех типов колбочек с разными спектральными чувствительностями. Спектральная чувствительность рецепторных потенциалов палочек примерно соответствует спектру поглощения родопсина с максимумом около 500 нм. У позвоночных рецепторный потенциал фоторецепторов в отличие от других рецепторов связан 250 ЧАСТЬ III. ОБЩАЯ И СПЕЦИАЛЬНАЯ СЕНСОРНАЯ ФИЗИОЛОГИЯ
не с деполяризацией, а с гиперполяризацией мембраны в ответ на адекватную стимуляцию (свет). Амплитуда (А) вторичного рецепторного потенциала коррелирует с интенсивностью светового стимула (Is) (т.е. с числом фотонов, падающих в единицу времени на единицу площади) следующим образом (рис. 11 .17, Б, В):
Эта гиперболическая функция («закон Геринга») в диапазоне промежуточных интенсивностей приближается к логарифмической, которая в общей сенсорной физиологии известна как закон ВебераФехнера (см. с. 188, 189):
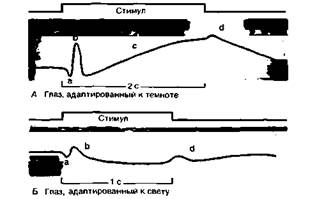
где I0- пороговая интенсивность стимула, зависящая от уровня адаптации. Константы а, к и к* в уравнениях (9) и (10) меняются вместе с длиной волны монохроматического света. Эта зависимость примерно соответствует кривым спектральной чувст- вительности рецепторных потенциалов (рис. 11.18) [8, 36]. Роговично-сетчаточный потенциал и электроретинограмма (ЭРГ) С помощью макроэлектродов от всего глаза можно отводить два функционально различных типа электрических потенциалов: постоянный между роговицей и более отрицательной сетчаткой - и электроретинограмму (ЭРГ). Первый обусловлен главным образом разностью потенциалов между склеральной стороной клеток пигментного эпителия и внутренними сегментами фоторецепторов. Другими словами, он отражает суммарный электрический ток, проходящий через плазматические мембраны пигментных клеток и фоторецепторов, поэтому меняется в зависимости от уровня адаптации сетчатки. ЭРГ-это колебания отводимого от всего глаза напряжения, вызываемые увеличением или уменьшением количества падающего на сетчатку света. ЭРГ включает несколько «волн»: а, b, с и d (рис. 11.19). Волна а возникает, по-видимому, за счет суммации рецепторных потенциалов; более медленная b-волна отражает изменения, главным ГЛАВА 11. ЗРЕНИЕ 251
образом мембранных потенциалов биполярных и глиальных (мюллеровых) клеток, с-волна - мембранных потенциалов клеток пигментного эпителия при «включении света» (on-эффект), а d-волна— мембранных потенциалов фоторецепторов и биполярных клеток при «выключении света» (off-эффект). Если длительность светового стимула превышает 0,3 с, с-волна появляется еще в ходе его действия и off-эффект накладывается на нее. При коротких световых вспышках b- и d-волны совпадают [8, 10]. ЭРГ обращаемого узора регистрируется при рассматривании узора, светлые и темные участки которого периодически меняют яркость на противоположную (например, при обращении цвета светлых и темных шахматных полей на телеэкране). Средняя яркость такого стимула неизменна, поэтому ЭРГ в данном случае генерируется в основном ганглиозными клетками сетчатки, т.е. нейронными элементами, особенно сильно реагирующими на изменения контраста и контрастные границы. Если ганглиозные клетки дегенерируют, ЭРГ обращаемого узора исчезает. Такой симптом в медицине обычно связан со слепотой или с резким ухудшением остроты зрения (см. с. 264).
|

 (9)
(9) (10)
(10)