
Развитие научных подходов, используемых в экологических исследованиях
В ходе развития экологии как опытно-экспериментальной сферы научной деятельности формировались и доминировали различные способы организации исследований [28, 33, 34, 35]. Для современной экологии характерно равноправное существование следующих подходов: 1) популяционный; 2) экосистемный; 3) синэкологический; 4) биотический анализ; 5) эволюционно-исторический; 6) комплексный, 7) социально-демографический; 8) геоинформационый и другие. Популяционный подход. Одной из важнейших задач популяционной экологии было выявление общих закономерностей динамики численности популяций — как отдельно взятых, так и взаимодействующих (например, конкурирующих за один ресурс или связанных отношениями «хищник—жертва»). Для решения этой задачи использовались простые математические модели — формулы, показывающие наиболее вероятные связи между отдельными, характеризующими состояние популяции величинами: рождаемостью, смертностью, скоростью роста, плотностью (числом особей на единицу пространства), и др. Математические модели позволяли проверять следствия разных допущений, выявив необходимые и достаточные условия для реализации того или иного варианта популяционной динамики. В 1920г. американский исследователь Р. Перль (1879-1940) выдвинул так называемую логистическую модель популяционного роста, предполагающую, что по мере увеличения плотности популяции скорость ее роста снижается, становясь равной нулю при достижении некоторой предельной плотности. Изменение численности популяции во времени описывалось S-образной кривой, выходящей на плато. Перль рассматривал логистическую модель как универсальный закон развития любой популяции. И хотя вскоре выяснилось, что это далеко не всегда так, сама идея о наличии некоторых основополагающих принципов, проявляющихся в динамике множества разных популяций, оказалась очень продуктивной. Внедрение в практику экологии математических моделей началось с работ Альфреда Лотки (1880-1949). Свой метод он сам называл «физической биологией» — попыткой упорядочить биологическое знание с помощью подходов, обычно применяемых в физике (в том числе — математических моделей). В качестве одного из возможных примеров он предложил простую модель, описывающую сопряженную динамику численности хищника и жертвы. Модель показала, что если вся смертность в популяции жертвы определяется хищником, а рождаемость хищника зависит только от обеспеченности его кормом (т. е. числа жертв), то численность и хищника, и жертвы совершает правильные колебания. Затем Лотка разработал модель конкурентных отношений, а также показал, что в популяции, увеличивающей свою численность по экспоненте, всегда устанавливается постоянная возрастная структура (т. е. соотношение долей особей разного возраста). Позднее им же были предложены методы расчета ряда важнейших демографических показателей. Примерно в эти же годы итальянский математик В. Вольтерра, независимо от мнения Лотки, разработал модель конкуренции двух видов за один ресурс и показал теоретически, что два вида, ограниченных в своем развитии одним ресурсом, не могут устойчиво сосуществовать — один вид неизбежно вытесняет другой. Теоретические исследования Лотки и Вольтерры заинтересовали молодого московского биолога Г. Ф. Гаузе. Он предложил свою, гораздо более понятную биологам, модификацию уравнений, описывающих динамику численности конкурирующих видов, и впервые осуществил экспериментальную проверку этих моделей на лабораторных культурах бактерий, дрожжей и простейших. Особенно удачными были опыты по конкуренции между разными видами инфузорий. Гаузе удалось показать, что виды могут сосуществовать только в том случае, если они ограничены разными факторами, или, иначе говоря, — если они занимают разные экологические ниши. Данное правило, получившее название «закона Гаузе», долгое время служило отправной точкой в обсуждении межвидовой конкуренции и ее роли в поддержании структуры экологических сообществ. Результаты работ Гаузе были опубликованы в ряде статей и книге «Борьба за существование» (1934), которая при содействии Перла вышла на английском языке в США. Книга эта имела громадное значение для дальнейшего развития теоретической и экспериментальной экологии. Она несколько раз переиздавалась и до сих пор часто цитируется в научной литературе. Изучение популяций происходило не только в лаборатории, но и непосредственно в полевой обстановке. Важную роль в определении общей направленности таких исследований сыграли работы английского эколога Чарлза Элтона (1900-1991), особенно его книга «Экология животных», опубликованная впервые в 1927, а потом не раз переиздававшаяся. Проблема динамики численности выдвигалась в этой книге как одна из центральных для всей экологии. Элтон обратил внимание на циклические колебания численности мелких грызунов, происходившие с периодом в 3-4 года, а, обработав многолетние данные о заготовке пушнины в Северной Америке, выяснил, что зайцы и рыси тоже демонстрируют циклические колебания, но пики численности наблюдаются примерно раз в 10 лет. Много внимания Элтон уделял изучению структуры сообществ (предполагая, что структура эта строго закономерна), а также цепям питания и так называемым «пирамидам чисел» — последовательному уменьшению численности организмов по мере перехода от нижних трофических уровней к более высоким — от растений к травоядным, а от травоядных к хищникам. Популяционный подход в экологии долгое время развивался преимущественно зоологами. Ботаники же больше исследовали сообщества, которые чаще всего трактовали как целостные и дискретные образования, между которыми довольно легко провести границы. Тем не менее, уже в 1920-е годы отдельные экологи высказывали «еретические» (для того времени) взгляды, согласно которым разные виды растений могут по-своему реагировать на определенные факторы внешней среды, а их распределение вовсе не обязательно должно совпадать с распределением других видов того же сообщества. Из этого следовало, что границы между разными сообществами могут быть весьма размытыми, а само выделение их условно. Наиболее четко такой, опережающий свое время, взгляд на растительное сообщество был развит российским экологом Л. Г. Раменским. В 1924 в небольшой статье (ставшей потом классической) он сформулировал основные положения нового подхода, подчеркнув, с одной стороны, экологическую индивидуальность растений, а с другой — «многомерность» (т. е. зависимость от многих факторов) и непрерывность всего растительного покрова. Неизменными Раменский считал только законы сочетаемости разных растений, которые и следовало изучать. В США совершенно независимые сходные взгляды, примерно в те же годы, развивал Генри Аллан Глисон (1882-1975). В его «индивидуалистической концепции», выдвинутой в качестве антитезы представлениям Клементса о сообществе как об аналоге организма, также подчеркивалась независимость распределения разных видов растений друг от друга и непрерывность растительного покрова. По-настоящему работы по изучению популяций растений развернулись только в 1950-х и даже 1960-х годах. В России бесспорным лидером этого направления был Тихон Александрович Работнов (1904-2000), а в Великобритании — Джон Харпер. Экосистемный подход. Термин «экосистема» был предложен в 1935 видным английским экологом-ботаником Артуром Тенсли (1871-1955) для обозначения естественного комплекса живых организмов и физической среды, в которой они обитают. Однако исследования, которые с полным основанием можно назвать экосистемными, начали проводиться значительно раньше, а бесспорными лидерами здесь были гидробиологи. Гидробиология, а особенно — лимнология с самого начала были комплексными науками, имевшими дело сразу со многими живыми организмами, и с их средой. Изучались при этом не только взаимодействия организмов, не только их зависимость от среды, но и, что не менее важно, — влияние самих организмов на физическую среду. Нередко объектом исследований для лимнологов был целый водоем, в котором физические, химические и биологические процессы теснейшим образом взаимосвязаны. Уже в самом начале 20-го века американский лимнолог Эдвард Бердж (1851-1950) с помощью строгих количественных методов изучает «дыхание озер» — сезонную динамику содержания в воде растворенного кислорода, которая зависит как от процессов перемешивания водной массы и диффузии кислорода из воздуха, так и от жизнедеятельности организмов. Существенно, что среди последних как производители кислорода (планктонные водоросли), так и его потребители (большинство бактерий и все животные). В 1930-х годах большие успехи в изучении круговорота вещества и трансформации энергии были достигнуты в Советской России на Косинской лимнологической станции под Москвой. Возглавлял станцию в это время Леонид Леонидович Россолимо (1894-1977), предложивший так называемый «балансовый подход», уделяющий основное внимание круговороту веществ и трансформации энергии. В рамках этого подхода начал свои исследования первичной продукции (т. е. создания автотрофами органического вещества) и Г. Г. Винберг, используя остроумный метод «темных и светлых склянок». Суть его в том, что о количестве образовавшегося при фотосинтезе органического вещества судят по количеству выделившегося кислорода. Спустя три года аналогичные измерения были осуществлены в США Г. А. Райли. Инициатором этих работ был Джордж Эвелин Хатчинсон (1903-1991), который своими собственными исследованиями, а также горячей поддержкой начинаний многих талантливых молодых ученых, оказал значительное влияние на развитие экологии не только в США, но и во всем мире. Перу Хатчинсона принадлежит «Трактат по лимнологии» — серия из четырех томов, представляющая собой самую полную в мире сводку по жизни озер. В 1942 в журнале «Эколоджи» вышла статья ученика Хатчинсона, молодого и, к сожалению, очень рано умершего эколога — Раймонда Линдемана (1915-1942), в которой была предложена общая схема трансформации энергии в экосистеме. В частности, было теоретически продемонстрировано, что при переходе энергии с одного трофического уровня на другой (от растений к травоядным животным, от травоядных — к хищникам) количество ее уменьшается и организмам каждого последующего уровня оказывается доступной только малая часть (не более 10%) от той энергии, что была в распоряжении организмов предыдущего уровня. Для самой возможности проведения экосистемных исследований очень важным было то, что при колоссальном разнообразии форм организмов, существующих в природе, число основных биохимических процессов, определяющих их жизнедеятельность (а следовательно — и число основных биогеохимических ролей!), весьма ограничено. Так, например, самые разные растения (и цианобактерии) осуществляют фотосинтез, при котором образуется органическое вещество и выделяется свободный кислород. А поскольку конечные продукты одинаковы, то можно суммировать результаты активности сразу большого числа организмов, например, всех планктонных водорослей в пруду, или всех растений в лесу, и таким образом оценить первичную продукцию пруда или леса. Ученые, стоявшие у истоков экосистемного подхода, хорошо это понимали, а разработанные ими представления легли в основу тех крупномасштабных исследований продуктивности разных экосистем, которые получили развитие в разных природных зонах уже в 1960-1970-х годах. К экосистемному подходу примыкает по своей методологии и изучение биосферы. Термин «биосфера» для обозначения области на поверхности нашей планеты, охваченной жизнью, был предложен в конце 19-го века австрийским геологом Эдуардом Зюссом (1831-1914). Однако в деталях представление о биосфере, как о системе биогеохимических циклов, основной движущей силой которых является активность живых организмов («живого вещества»), было разработано уже в 1920-30-х годах российским ученым Владимиром Ивановичем Вернадским (1863-1945). Что касается непосредственных оценок этих процессов, то их получение и постоянное уточнение развернулось только во второй половине 20-го века, и продолжается до сих пор. Синэкологический подход. Сущность синэкологического подхода заключается в изучении биотических компонентов экосистемы. Основными принципами являются: 1) поэтапность сукцессионных изменений; 2) сбалансированность взаимосвязей в климаксовых сообществах. Данный подход используется для решения вопросов рационального использования природных ресурсов. Изучение мест обитания или биотический анализ. Сущностью биотического анализа является изучение участков среды определенного типа (местообитание, физическое пространство), а также необходимые для жизнедеятельности живых сообществ природные ресурсы (экологическая ниша) и частные территории (микро местообитания). Данный подход включает принципы следующих методов: А Методы по оценке размеров популяций: объективные и субъективные 1. Объективные методы: прямые и косвенные. К прямым методам относят: - метод квадрата (определение плотности или частоты проективного покрытия); - метод прямого наблюдения; - метод фотографирования; - метод прямого учета; - метод сбора организмов или образцов. К косвенным методам относят: - метод повторного отлова и мечения (определение индекса Линкольна); - метод изъятия. 2. Субъективные методы определяют обилие и покрытие популяции или экосистемы по стандартным шкалам. Б Изучение растительных ассоциаций (группы, сообщества) с доминирующим одним или двумя видами (например: ковыльно-полынная ассоциация характерна для Оренбургских степей). В Метод индикационных исследований (например: при изучении лишайников - определение объёма кислорода). Г Этологический метод (изучение поведения животных). Эволюционно-исторический подход. Сущность подхода заключается в изучении изменений связанных с развитием жизни и истории цивилизации, позволяющих понять закономерности, действовавшие до появления человека и после этого события. Данный подход включает в себя принципы методов геологических исследований и математического моделирования исторического развития экологических ситуаций в биосфере. Его значение состоит в следующем: 1. Долгосрочный прогноз возможных биосферных изменений, позволяющих сделать выводы о характере вероятных будущих преобразований в биосфере. 2. Изучение соотношения видов в ископаемых сообществах. 3. Изучение реликтов и эндемиков (гинго и тисс ягодный). Ранее этими вопросами занималась палеонтология и археология, но с дополнением экологического содержания появилась возможность изучать закономерности изменения климата и закономерности конвергентной коэволюции. Комплексный подход. Сущность подхода заключается в разработке критериев оценки состояния компонентов природно-техногенной системы. В настоящее время в практике оценочных исследований в качестве признаков применяют нормативные санитарно-гигиенические показатели и комплексные экологические критерии. Одни из таких индикаторов состояния экологических систем выступает суммарный показатель (Xs), рассчитываемый по формуле:
где n – число компонентных критериев; Хi – показатель компонента (в относительных величинах); Кi – массовый коэффициент показателя. Геоинформационный подход. Это система новых взглядов на окружающий нас мир, опирающийся на современные компьютерные технологии для картирования и анализа объектов реального мира, а также событий, происходящих на нашей планете. Этот подход объединяет основы прикладной информатики в области традиционных операций работы с базами данных, такими как запрос и статистический анализ, принципы экологической статистики с преимуществами полноценной визуализации и географического (пространственного) анализа, которые предоставляет карта [27, 32, 33, 34]. Данный подход позволяет формировать эколого-информационный портрет регионов и разрабатывать системы мер по принятию экологически грамотных решений специалистами в области защиты окружающей среды. Социально-демографический подход. Система методов оценки и прогноза развития изменений в состоянии здоровья социальных групп населения по различным показателям. Степень зависимости общей заболеваемости от качества среды
где Pmax – условная максимальная заболеваемость (100 % заболеваемость из-за загрязнения среды); а, b – параметры логистической функции; К – общая загрязненность среды.
|
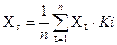 (1.1)
(1.1)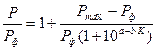 (1.2)
(1.2) - отношение общей заболеваемости к фоновой заболеваемости при отсутствии экопатологий, т.е. заболеваний, вызванных неблагоприятными экологическими факторами;
- отношение общей заболеваемости к фоновой заболеваемости при отсутствии экопатологий, т.е. заболеваний, вызванных неблагоприятными экологическими факторами;


