
Экологическую неравноценность местообитаний сурков.
Байбак. Неравноценность местообитаний восновном определяют возможности кормежки (продолжительность и сроки вегетации трав) и в меньшей степени защитность и условия норения. Наши наблюдения подтверждают наблюдения многих исследователей (Шубин, 1963; Капитонов, 1969; Бибиков, 1967, 1989 и др.) о влиянии на выживаемость зверьков и успешность их воспроизводства осадков и температуры окружающей среды. Засушливость климата препятствует широкому распространению байбаков в обширных зонах пустынных степей и полупустынь (Шубин и др., 1978), так как из-за раннего выгорания растительности сурки не успевают набрать необходимые для зимней спячки запасы жира. Помимо засушливости на численности молодых особей отрицательно сказываются длительные весенние или поздневесенние возвратные холода, задерживающие вегетацию растительности. Таблица 3.1.2 Площадь семейного участка (га) сурка Мензбира и серого сурка
* над чертой пределы, под чертой среднее, ** на платообразных всхолмленных участках, *** по берегам рек
После сухого лета сеголетки залегают в спячку, не имея достаточного количества жира и после зимовки годовалых зверьков практически не остается (Шубин и др., 1978). Но и избыток осадков не всегда благоприятно влияет на сурков. Из-за пониженной наземной активности звери медленнее набирают жир и к спячке накапливают его даже меньше, чем при жаркой погоде с умеренной нормой осадков. В основной массе байбаки спариваются в норах до выхода на поверхность после спячки, поэтому погода не может оказать влияние на протекание беременности. Более важным в жизни самок является период лактации, приходящийся на раннюю весну. Если в это время стоит теплая погода, способствующая нормальному развитию в местообитаниях сурков растительности, самки могут ежедневно питаться и сытно кормить молоком своих малышей. Задержка развития растительности из-за холодной ненастной погоды весной приводит к истощению лактирующих самок и постэмбриональной гибели молодняка. Таким образом, основным лимитирующим фактором, обеспечивающим благополучие прироста у байбака в Казахстане, является теплая погода ранней весной и перед залеганием в спячку. Сохранность молодняка в период лактации в европейской части страны выше, так как рождение детенышей происходит в период обилия кормов (Середнева, 1975; наши данные)). При благоприятных кормовых условиях до годовалого возраста доживает до 57% приплода, а при неблагоприятных - почти весь приплод погибает (Середнева, 1991).Мы считаем это заявление спорным, поскольку за период более 30 лет мы не отмечали 100% гибели молодняка. Больше всего зверьки гибнут во время первой зимней спячки, особенно после засушливого лета, что отмечалось в восточных районах Оренбургской области в засушливое жаркое лето 2010 года. Мало набравшие жира сеголетки почти все погибли во время зимовки. От числа ушедших в зимовку зверьков весной на поверхность вышло лишь 11% особе 2010 года рождения (Охотовед Ахтямов Ф., личное сообщение). Все отмеченные колебания происходят в пределах естественного саморегулирования в популяциях (если не учитывать антропогенный фактор). В норме у байбаков в Казахстане ежегодно приплод отмечается в 28,6-40,8, в среднем 32,9% семей. При среднем количестве малышей в выводке 4,3 и средней численности в семье 4,8 особей такого воспроизводства достаточно для поддержания неопромышляемой популяции в равновесном состоянии (Машкин, 1988). В европейской части страны в охраняемых популяциях с выводками бывает 26,6-30,8% семей (Тихвинский, 1934), 37,5-61,1% (наши данные). Размеры смертности во всех частях ареала байбака примерно одинаковы, в Казахстане она составляет 33,9% от общей численности, в том числе, сеголетки - 15,2%. До годовалого возраста от числа появившихся на поверхности сеголетков доживает 31,9-62,8, в среднем 52,4% особей. Прирост поголовья в неопромышляемых популяциях составляет 40-47%. Усиленное размножение отмечается в эксплуатируемых популяциях, как компенсаторная реакция на убыль. Прирост в опромышляемых поселениях достигает 115-125%. В отдельные годы в некоторых местах - 26,6 или 225%, а в среднем 44,9-51,5 (Шубин, 1967). Таким образом, при прогнозировании численности байбака следует основываться на метеорологических данных и материалах о ходе вегетации растений в местообитаниях сурков в период 1-1,5 месяца после выхода зверей из спячки и в последний месяц перед уходом в зимнюю спячку. Универсальным показателем, характеризующим состояние популяции, является прирост поголовья (Р), который рассчитывается по данным учета особей в семьях (Машкин, 1986) по формуле 1: Р = С/М · 100%. (1), где Р - прирост популяции, %;С - численность сеголетков; М - численность сурков в возрасте 1 года и старше. Численность особей в семьях по годам существенно меняется в результате рождаемости, смертности, эмиграции и иммиграции половозрелых зверей (перераспределение в семьях). Значительные изменения в состав вносит рациональный промысел. Но ни один из перечисленных факторов не ведет к уничтожению семей. Для получения приемлемых для практики результатов необходимо обследовать при учетных работах минимальную выборку в 30-40 семей. То есть, подсчитав в 30-40 семьях число выводков, сеголетков и общее число особей можно по формуле (1) достаточно объективно определить прирост изучаемой популяции. Такой объем выборки необходимо сделать для местообитаний с низкой плотностью населения сурков (1-10 семей на 1 км²), а затем такое же количество обследовать в поселениях со средней (11-30) и с высокой плотностью (31 и более семей на 1 км²). Наши исследования показали, что если выводки имеются менее, чем в 30% семей и прирост популяции составляет менее 35%, то следует ставить вопрос о немедленном прекращении промысла, выяснении причин снижения воспроизводства и последующем наблюдении за состоянием популяции. Низкий прирост свидетельствует о подрыве воспроизводственного поголовья, или очень неблагоприятном ходе размножения, или слабой выживаемости молодняка. Серый и красный сурки. У горных видов из-за расчлененного рельефа очень велика мозаичность местообитаний, поэтому там четко выделяют три типа поселений: диффузный (степной), ленточный (балочный) и пятнистый (очаговый) (Бибиков, 1967). Наибольшая емкость угодий в поселениях степного типа. Благодаря однообразию растительности, резких сезонных изменений в использовании территории участка не наблюдается. Но в отдельные неблагоприятные годы с влажной холодной весной у половины размножавшихся самок рассасывается до 25% эмбрионов, и смертность сурчат в течение первого года жизни достигает 70-81% (Михайлюта, Кулькова, 1969). При ленточном типе емкость угодий ниже, поэтому в пределах площади пригодных местообитаний около половины, а нередко и большая часть территории, не имеет нор. Более сложная топография и мозаичность растительности в ленточных поселениях создает благоприятные условия для переживания весенних холодов и летних засух, позволяя зверькам перемещаться в пределах семейного участка и сменять пастбища. При очаговом типе поселений заселенная площадь не превышает 10% территории, пригодной для жизни сурков. Здесь зверьки летом широко рассредотачиваются по прилегающим малопригодным для зимовки территориям (Бибиков, Берендяев, 1978). Оценивая изменчивость плотности населения горных видов сурков, было выявлено наличие у них устойчиво населенных стаций переживания («ядер поселения»), то есть установлена экологическая неравноценность отдельных частей их поселений (Бибиков, Жирнова, 1956; Чекалин, 1965; Жаров, 1972; Сунцов, 1981). Менее благоприятные для жизни участки поселения обычно имеют невысокую и колеблющуюся по годам численность зверей. В благоприятные годы в экологически неустойчивых поселениях численность сурков может увеличиваться в несколько раз без увеличения внешнего контура и площади поселения, в оптимальных же население растет незначительно (Бибиков и др., 1973). Оптимальные местообитания приурочены обычно к террасам ручьев, верховьям долин. Менее благоприятные для жизни участки примыкают к оптимальным, располагаясь на склонах и по днищам долин. При снижении численности населения эти местообитания часто лишены сурков. Без применения метода картирования при выявлении экологической неравноценности отдельных частей поселений нельзя установить наличие значительных изменений численности сурков, поэтому для горных видов особенно актуально стоит вопрос о проведении регулярных учетов их численности в семьях и картировании общих контуров поселений. Для горных сурков показателем роста или снижения численности, учитывающем естественный отход или промысловое изъятие, является показатель прироста. В норме у серых сурков в размножении участвует 30-38% взрослых самок (Деревщиков, 1967). В другой части ареала при возрастании плотности населения доля размножающихся самок снижается с 70 до 47%, а размер выводка уменьшается с 6,4 до 5,0 особей (Поле, 1970). Поскольку у горных сурков смертность молодняка в среднем несколько выше, а половой зрелости большинство самок достигает позднее, чем у равнинного байбака, то критический прирост, обеспечивающий воспроизводство популяции следует считать 40-45%. В отдельные годы в норме прирост колеблется от 54,7 до 229% (Бибиков, Берендяев, 1978; Кизилов, Берендяев, 1978). Тарбаган. По характеру размещения по территории, строению нор, темпам размножения тарбаганы ближе к байбакам, чем к горным видам сурков. Число самок, приносящих приплод, обычно не превышает 50%, но в отдельные годы может колебаться от 17 до 77% (Летов и др., 1978). Прирост поголовья при среднем выводке 4,5 особи, но при различной доле участия самок в размножении, колеблется в пределах 20-162%. В норме для поддержания гомеостатического равновесия популяции ежегодный прирост должен составлять не менее 35-40%. Сурок Мензбира. Отличительной чертой района обитания сурка Мензбира является большое количество выпадающих зимой и весной осадков и почти полное их отсутствие летом. Поэтому все поселения приурочены к болотистым участкам (сазам), снежникам, долинам рек и ручьев. При сезонных перемещениях в пределах семейного участка звери даже в самые засушливые годы не страдают от бескормицы. То есть условия жизни сурка Мензбира достаточно стабильны, и погодные условия не влияют на изменения численности. Смертность молодняка самая низкая среди всех рассматриваемых видов сурков. До годовалого возраста от числа впервые вышедших на поверхность сеголетков доживает 51-76%, в среднем 67%. Но у этого вида самая низкая плодовитость. Выводки состоят из 1-5, в среднем, 2,8 особей. Ежегодно в норме выводки отмечаются в 32,4-62,6, в среднем у 39,7% семей (Машкин, 1989), то есть ежегодный прирост составляет 17,8-41,4, в среднем 29,9%. Наблюдения показали, что при участии 30% самок в размножении прирост не восполняет убыль. Критический прирост около 18-20%. Черношапочный сурок. Распространен этот зверь спорадически и численность его невелика. Климат в его местообитаниях отличается суровостью. Важнейшим фактором, определяющим размещение и численность сурков, является снежный покров, спасающий сурчиные жилища от зимнего промерзания. Летом, на постепенно вытаивающих из под огромных снежных надувов участков, зверьки обеспечены свежевегетирующей растительностью до спячки. У черношапочного сурка самые многочисленные выводки, состоящие из 3-11, в среднем 6 малышей, в северных районах обитания (Капитонов, 1963) и 1-7, в среднем, 4,6 особей, в южных районах (Жаров,1972). Потомство ежегодно приносят до 69-76% самок. Учитывая постоянство кормовой базы и ежегодную хорошую упитанность зверей перед спячкой, В. И. Капитонов (1963) и В. Н. Жаров (1970) считают, что эти показатели участия самок в размножении из года в год довольно постоянные. Неблагоприятные условия норения черношапочных сурков в вечномерзлых грунтах приводят к высокой смертности зимой. Среди молодых зверьков численность после спячки сокращается на 50-58%, а среди всех зверей - на 33% (Капитонов, 1978). Из приведенных данных можно заключить, что прирост в популяциях черношапочного сурка в северных районах обитания достигает 317%, а в южных - 142%. Самые высокие приросты поголовья в нормальных популяциях черношапочного сурка обусловлены резко континентальным климатом. Поэтому, видимо, и минимально допустимый прирост не должен быть менее 70%. Перечисленные в таблице 3.1.3 показатели помогут обосновать краткосрочный прогноз, а показатель прироста популяции является интегрированным методом оценки состояния популяции сурков. Следует помнить, что в зависимости от конкретных погодных условий года и состояния хозяйственного воздействия на популяцию прирост и доля семей с выводками могут несколько отклоняться от приведенных в таблице 3.1.3 средних многолетних данных. Таблица 3.1.3 Экспресс-оценка состояния популяции сурков по показателям прироста поголовья у разных видов
Дополнительными критериями оценки состояния популяции сурков являются лимитирующие экологические факторы, которые проявляются в приведенных в таблице 3.1.4 признаках. Связь сроков залегания в спячку с выпасом КРС. В течение многолетних наблюдений отмечена растянутость сроков залегания сурков в спячку (табл. 3.1.4), обусловленные разнокачественностью кормовых ресурсов участков обитания. Таблица 3.1.4 Экологическая оценка состояния популяции сурков
Залеганию в спячку предшествует период, который характеризуется слабой вегетацией растительности. Первыми (в первой половине сентября) залегают в спячку животные, чьи участки находятся на территории без выпаса КРС. Сроки залегания этих семей относительно постоянны и не зависят от погодных условий. Залеганию в спячку предшествует период, который характеризуется усыханием растительности. Последними закрываются норы на участках с выпасом КРС, состояние кормовых ресурсов которых ранее (Ронкин, Савченко, 1993, 1996, 1999) было признано как наиболее благоприятное. В годы с сухой осенью сроки залегания в спячку более сжаты (с начала по конец октября), чем в годы с влажной осенью (с начала октября по конец ноября). Изменение кормовых условий на одном и том же участке влечет за собой соответствующие изменения сроков залегания в спячку. Залеганию в спячку сурков в местообитаниях любого типа предшествует не менее чем 2-недельный период неблагоприятных кормовых условий, который, вероятно, может служить сигналом приближающегося начала спячки. И даже если этот период сменяется периодом усиленной вегетации, сурки все равно залегают в спячку. В случае стабильных кормовых условий (при равномерно влажной осени), период активности отдельных особей может продолжаться до 3-ей декады ноября. Сроки залегания в спячку могут служить критерием благоприятности кормовых условий. Динамика залегания в спячку отражает соотношение числа семейных участков, находящихся в тех или иных кормовых условиях, и насколько участки разнятся по этим условиям. Типы поселений. Совокупности семей образуют более крупные территориальные структуры - колонии, занимающие сравнительно обособленные территории и объединенные зрительно-звуковой связью отдельных семей. Степень обособленности колоний зависит от ландшафтных особенностей местообитаний, которые определяют тип поселения. При мозаичном, или очаговом, типе поселения группы семей легко вычленяются. В балочных поселениях колонии протягиваются на значительные расстояния вдоль склонов оврагов, ущелий, русел речек и пр. Разрывы в непрерывной ленте семей, обусловленные какими-либо препятствиями (выходы скал, заболоченные или распаханные участки, лесонасаждения и пр.), из-за которых между крайними семьями нарушается зрительно-звуковая связь, являются естественными разделителями отдельных колоний. Обычно колонии достаточно автономны, но через расселение достигших половой зрелости особей они функционально связаны с другими существующими или образующимися колониями. Если биологические характеристики сурков отвечают биотопическим параметрам ландшафта, в котором они обитают или который заселяют вновь, то такие колонии жизнеспособны и могут существовать бесконечно долго. Когда семьи сурков размещены на обширных территориях диффузно и нигде характер межсемейных связей не нарушается, в этом случае уместно говорить о поселениях - более крупных территориальных структурах. Поселение - это совокупность функционально взаимосвязанных отдельных семей или колоний, занимающих определенный тип ландшафта, в котором обитают сурки. Выделяют три типа поселений: степной, или диффузный, балочный, или ленточный, и мозаичный, или очаговый (Бибиков, 1967;). Степные поселения. Отмечаются в относительно выровненных местообитаниях высокогорных сыртовых поясов, платообразных возвышенностей, террасированных склонов рек и обширных степных равнин. Семьи сурков в этих поселениях размещены относительно равномерно, и поселения тянутся на многие десятки километров. Это, как правило, наиболее “древние” монолитные поселения с хорошо выраженными крупными бутанами, с системой временных защитных нор. Сурчины распределены довольно равномерно, незаселенных мест почти нет. Плотность населения зверьков на гнездопригодных участках обычно средняя и высокая. Однообразие растительности определяет сравнительно крупные семейные участки. Этот тип поселений характерен для байбака казахстанского, обитающего в местах, где сохранились большие целинные массивы. Вместе с тем внутри “клеток” пахотных массивов остались небольшие участки целины, и сурки живут на них и на прилегающих кромках полей. Особенно благоприятные условия складываются на полях со сменой выращиваемых культур (сеяные травы, зерновые, технические культуры). На таких массивах формируются поселения, отличающиеся от исходных поселений степного типа. Они свойственны всем основным типам сельскохозяйственных угодий и сохраняют структуру при разных уровнях численности. Наиболее стабильные “ядра” приурочены к краям пахотных “клеток”. Такой тип поселений назван “сетчатым” (Румянцев, 1991). Балочный тип. Преобладает в расчлененном рельефе гор, по оврагам, балкам и склонам речек на относительно равнинных территориях; характерен для большей части местообитаний горных видов сурков и европейского байбака, норы которого располагаются по склонам балок, промоин, вдоль русел речек и ручьев, протягиваясь на значительные расстояния по водосборной сети. При сглаженном рельефе по вершинам ручьев ленты различных поселений могут соединяться. На обширных склонах семьи располагаются в несколько ярусов и, как правило, в нижележащих семьях размер семейного участка меньше, чем у крайних верхних. На выровненных участках бутаны крупные, чем на склонах, где выбрасываемый грунт смывается водой. В балочных поселениях очень велика мозаика растительных ассоциаций, разнообразны грунтовые условия, поэтому разные части семейных участков неравнозначны. В засушливые годы в степных поселениях сурки могут оказаться в трудных условиях, не набирают к спячке необходимое количество жира, в таких случаях отмечается повышенный отход зверей, особенно молодых (Страутман, 1956; Шубин и др., 1978). В балочных поселениях из-за различной экспозиции склонов и разнообразия фенологических фаз развития растений, приуроченности нор к снежным надувам, ручьям сурки оказываются в более благоприятных условиях и легче переносят климатические катаклизмы. Характер расположения семей предполагает лучшие возможности расселения мигрантов, нежели в степных поселениях, и определяет направление миграций. Плотность населения в балочных поселениях преимущественно средняя, редко высокая и низкая, что обусловлено пестротой рельефа, наличием каменистых осыпей, морен и различных неудобий, непригодных для жизни сурков. Очаговый тип поселений. Преобладает на периферии ареала, у верхнего или нижнего предела распространения в горах и на участках, где часть территории не обеспечивает основные жизненные потребности зверьков. Семьи распределены в угодьях одиночно или разрозненными группами. Этот тип поселения характеризуется наименьшей плотностью сурков, в сравнении со степным и балочным, но и здесь могут быть мозаично расположенные и разобщенные естественными преградами колонии со средней и даже высокой плотностью семей.
|
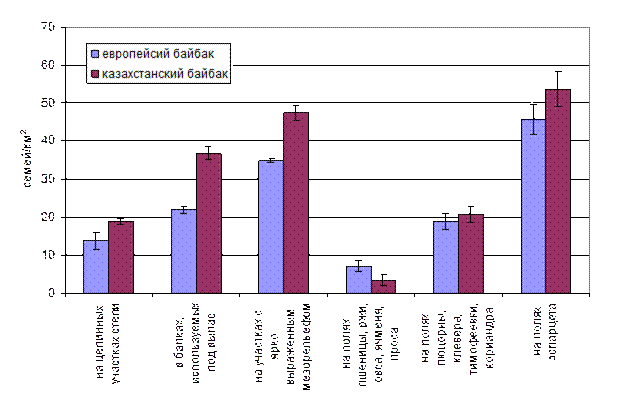 Рис.3.1.1 Средние плотности популяции байбаков в различных местообитаниях
Рис.3.1.1 Средние плотности популяции байбаков в различных местообитаниях


